目次

放射状グリア細胞(Radial Glial Cell)は、私たちの大脳皮質をつくるすべての神経細胞(ニューロン)とグリア細胞を生み出す「神経幹細胞」であると同時に、生まれたばかりのニューロンを正しい場所まで導く「足場(ガイドレール)」という、二つの大きな顔を持つ特別な細胞です。かつては単なる「配線ケーブル」と誤解されていましたが、いまでは脳づくり全体を統括する主役として理解されています。この記事では、放射状グリア細胞の働きと、その不具合が小頭症や滑脳症などの皮質形成異常にどうつながるのかを、遺伝専門医の視点でわかりやすく解説します。
Q. 放射状グリア細胞とは何ですか?まず結論だけ知りたいです
A. 放射状グリア細胞は、胎児期の脳で「神経幹細胞(大もとの細胞)」として働き、大脳皮質のニューロンとグリアを生み出す一次前駆細胞です。同時に、脳の内側から外側へ長い突起を伸ばし、新しく生まれたニューロンが目的の層までよじ登るための「足場」にもなります。この二つの役割のどこかが遺伝子の変化でうまく働かなくなると、小頭症・滑脳症・敷石状滑脳症・脳室周囲結節性異所性灰白質といった皮質形成異常が起こります。
- ➤正体 → 脳の最も内側(脳室帯)にあり、皮質のニューロンとグリアを生み出す神経幹細胞
- ➤二つの顔 → ①細胞を生み出す「幹細胞」 ②ニューロンを導く「足場」
- ➤ヒトらしさの鍵 → 外側放射状グリア(oRG)の増加が、大きく複雑なヒトの脳をつくった
- ➤関わる病気 → 小頭症・滑脳症・敷石状滑脳症・脳室周囲結節性異所性灰白質
- ➤遺伝診療との接点 → 原因遺伝子の同定はNGSパネル検査で行い、遺伝カウンセリングにつながる
1. 放射状グリア細胞とは?まず結論から
放射状グリア細胞は、胎児の脳の中で「大もとの細胞」として働く神経幹細胞です。名前に「グリア」とついているため補助的な細胞のように聞こえますが、実際には大脳皮質を構成するほとんどすべてのニューロンとグリア細胞を生み出す、脳づくりの中心的な設計者です。細胞のかたちは非常に特徴的で、脳の最も内側にある「脳室帯」に細胞の本体を置きながら、脳の表面(軟膜)に向かって1本の長い突起をまっすぐ放射状に伸ばしています。この「放射状に伸びる」姿こそが、名前の由来になっています。
この細胞がすごいのは、「細胞を生み出す工場」であると同時に、「生まれた細胞を運ぶ道路」という二役を一手に引き受けている点です。工場としては分裂を繰り返して新しいニューロンをつくり、道路としてはそのニューロンが目的地までよじ登っていくための足場になります。脳という複雑な立体構造を、細胞一つひとつが迷子にならずに組み上げていくために、この二役は欠かせません。
💡 用語解説:神経幹細胞と前駆細胞
神経幹細胞とは、自分と同じ細胞をつくる「自己複製」の能力と、ニューロンやグリアなど複数の種類に変化できる「多分化能」の両方を持つ、脳づくりの大もとの細胞です。ここから生まれ、もう少し役割が絞られた細胞を前駆細胞と呼びます。放射状グリア細胞は、皮質発生のピーク時に増えている細胞のほとんどを占める「一次前駆細胞(もっとも上流の幹細胞)」にあたります。
なぜこの基礎的な細胞の話を、遺伝医療のサイトで取り上げるのでしょうか。それは、放射状グリア細胞の働きが遺伝子の変化でうまくいかなくなると、脳の大きさや形そのものに関わる重い病気(小頭症や滑脳症など)につながるからです。これらの診断では、原因となる遺伝子をNGSパネル検査で調べ、その結果をもとに遺伝カウンセリングを行います。つまり放射状グリア細胞は、基礎生物学と臨床の遺伝診療をつなぐ「結び目」のような存在なのです。
2. 「ただのケーブル」からマスターセルへ:発見の歴史
放射状グリア細胞が最初に記録されたのは1885年、銀染色法(ゴルジ染色)を発明したカミッロ・ゴルジの手によるものでした。ゴルジは胚のニワトリの脊髄で、中心の管から外側へ放射状に長い突起を伸ばす特徴的な細胞を描き出しました。しかしその後の約100年間、この細胞の役割は大きく誤解され続けます。当時の定説では、ニューロンをつくる細胞とグリアをつくる細胞は完全に別系統だと考えられ、放射状グリア細胞は「新しいニューロンが皮質へ移動するときの受動的な足場(ケーブル)」にすぎず、役目を終えたらアストロサイトに変わって消えていく脇役だとみなされていました[1]。
この固定観念が覆ったのは2000年前後です。神経発生のピーク時に増えている前駆細胞のほとんどが、放射状グリア細胞の目印(GLAST・BLBP・RC2など)を持つことが偶然わかり、さらに特定の細胞だけを蛍光で標識して取り出す実験によって、長い突起を持つこの細胞集団こそがニューロン・グリア・幹細胞の大部分を含むことが証明されました[2]。決定打となったのは2001年、生きた脳のスライスをリアルタイムで観察する技術によって、放射状グリア細胞が実際に分裂し、そこからニューロンが生まれる瞬間が直接とらえられたことです[1]。こうして放射状グリア細胞は「単なる通り道」から「脳の設計を司るマスターセル」へと評価が一変しました。
🔍 関連記事:ネスチン(神経幹細胞マーカー)/ビメンチン/GFAP遺伝子
3. 細胞のかたちと「エレベーター運動」
放射状グリア細胞は、神経管ができた直後に存在する「神経上皮細胞」から生まれます。もっとも印象的な特徴が、細胞の核が上下に移動するエレベーター運動です。脳室帯にぎっしり詰まった細胞たちは、DNAをコピーする時期(S期)には核を脳室面から離れた外側へ移動させ、いよいよ分裂する時期(M期)になると核を脳室面へ一気に下降させます。まるでエレベーターのように核が行き来するため、この名で呼ばれます。
💡 用語解説:エレベーター運動(IKNM)
正式には核間分裂運動(Interkinetic Nuclear Migration:IKNM)といいます。細胞周期の進み具合に合わせて核が上下に移動する現象で、狭い脳室帯にたくさんの細胞を効率よく詰め込み、同時並行でどんどん分裂させるための工夫だと考えられています。詳しくはエレベーター運動(IKNM)の解説ページもご覧ください。
重要なのは、放射状グリア細胞が分裂している最中でも、軟膜へ向かう長い突起を切り離さずに保ち続けることです。この突起は後からやってくるニューロンの通り道になるため、途中で切れてしまうと道が失われてしまいます。分裂しながらも道路を維持し続ける——この几帳面さが、整然とした皮質の層構造を可能にしています。
分裂を終えた放射状グリア細胞は、直接ニューロンを生み出すこともあれば、いったん中間前駆細胞という「増幅役」の細胞を経由することもあります。中間前駆細胞は長い突起を捨てて脳室下帯へ移動し、そこで対称分裂を繰り返して一気に複数のニューロンを量産します。この間接的なつくり方(間接的神経産生)は、ニューロンの数を爆発的に増やす仕組みであり、とりわけ哺乳類の大きな大脳皮質を用意するうえで重要な役割を果たしています[1]。
4. 何を目印に見分けるのか:分子マーカー
放射状グリア細胞は、神経上皮細胞の未熟さと、成熟したアストロサイトに似た性質をあわせ持つ「移行期」の細胞です。そのため、研究者はさまざまな目印(マーカー)を組み合わせてこの細胞を見分けます。細胞の骨組みをつくるタンパク質では、未分化な状態を示すネスチンやビメンチンが強く現れ、発生が進むにつれてアストロサイトの代表的マーカーであるGFAPへと置き換わっていきます。
細胞の運命を指揮する「転写因子」も重要な目印です。神経板の段階から現れるSOX2は、未分化な増殖状態を保つ中心的な因子です。さらにPAX6は、神経上皮細胞から放射状グリア細胞への移行を促し、皮質ニューロンをつくるペースを調整する司令塔として機能します。Notchシグナルの下流で働くHES1・HES5は、細胞が早まって分化してしまわないようブレーキをかけ、幹細胞のプールを守っています。
💡 用語解説:転写因子と単一細胞RNAシーケンス
転写因子とは、どの遺伝子を「読む・読まない」を切り替えるスイッチ役のタンパク質で、細胞の性格を決めます。単一細胞RNAシーケンス(scRNA-seq)は、細胞を1個ずつバラバラにして、それぞれがどの遺伝子を使っているかを網羅的に読み取る技術です。この技術のおかげで、これまで均一に見えていた放射状グリア細胞の中に、実は少しずつ性質の違う「サブタイプ」が隠れていることが見えるようになりました。
実際、ヒトの発生期の脳で500個以上の前駆細胞を1個ずつ調べた研究では、PAX6・SOX2・GLAST・BLBP・ビメンチンがほぼすべての細胞(約93%)で検出され、放射状グリアとしての共通の土台が確認されました[3]。一方で、細かく分類すると、これらの細胞は複数の異なる状態に分かれることも示されました。とくにヒトに多いサブタイプでは、HOPX・PDGFD・ITGB5といった新しい目印が見つかっており、放射状グリア細胞が「均一な集団」ではなく、状況に応じて遺伝子のスイッチを動的に切り替える不均一な集団であることがわかっています[3]。
5. 対称分裂と非対称分裂:脳の細胞数を決めるスイッチ
最終的に脳のニューロンが何個になるかは、放射状グリア細胞が行う「対称分裂」と「非対称分裂」の切り替えで決まります。発生の初期には、母細胞から同じ性質の放射状グリア細胞が2個できる「対称分裂」が中心で、幹細胞のプールをねずみ算式に増やします。プールが十分に育つと、今度は「非対称分裂」が主役になります。これは、片方は幹細胞のまま残し、もう片方はニューロン(または中間前駆細胞)へと分化させる、性質の異なる2個の娘細胞を生む分裂です。
💡 用語解説:対称分裂と非対称分裂
対称分裂は、まったく同じ性質の細胞を2個つくる分裂で、幹細胞の「数を増やす」ときに使われます。非対称分裂は、幹細胞1個と、分化していく細胞1個という「別々の運命の細胞」を1回で生み出す分裂です。このスイッチのタイミングが早すぎると幹細胞が枯れてニューロンが足りず、遅すぎると分化が進まない——そのバランスが脳の最終的な大きさを左右します。
非対称分裂で運命を分ける鍵は、細胞の脳室側(頂端側)に集まるPar3/aPKC極性複合体という「目印タンパク質」の分配です。分裂のときの割れ目(分裂面)がこの目印をどう二分するかで、娘細胞の運命が決まります[4]。Par3を多く受け継いだ娘細胞ではNotchシグナルが強く入り、分化にブレーキがかかって幹細胞のまま残ります。反対にPar3が少なかった娘細胞ではNumbというタンパク質が働いてNotchが弱まり、ニューロンや前駆細胞への道を進みます[5]。この繊細なバランスを実験的に崩すと、娘細胞の運命指定が乱れることが確かめられています[5]。
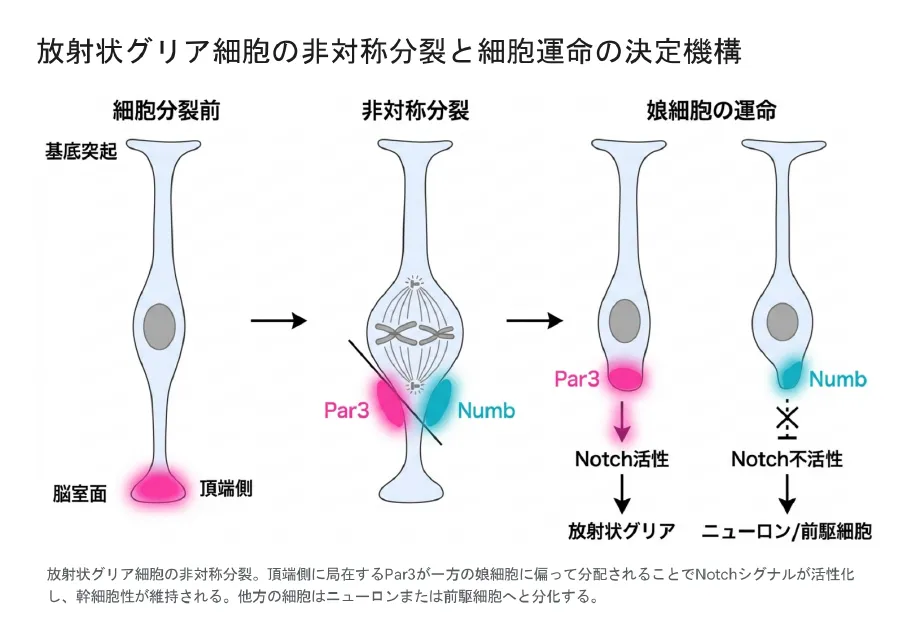
非対称分裂のしくみ。頂端側に集まるPar3が片方の娘細胞に偏って分配されるとNotchシグナルが活性化し、幹細胞性が保たれる。もう片方の細胞はNumbの働きでNotchが弱まり、ニューロンや前駆細胞へと分化する。
6. 外側放射状グリア(oRG)とヒトの脳の進化
ヒトや霊長類の大脳皮質は、マウスなどの齧歯類に比べて圧倒的に大きく、脳の表面に複雑なシワ(脳回)を持ちます。この「脳の巨大化」の原動力として近年注目されているのが、外側放射状グリア(oRG/基底放射状グリア)という特別な幹細胞の増加です。ヒトの脳では、脳室下帯が外側へ大きく厚みを増し、「外側脳室下帯(OSVZ)」という広大な増殖の場が生まれます。ここにびっしり存在するのがoRGです[6]。
💡 用語解説:外側放射状グリア(oRG)
古典的な放射状グリア(脳室側にある「vRG」)とちがい、oRGは脳室面との接触(頂端突起)を失っています。つまり上皮としての「アンカー」を持たない自由な細胞ですが、軟膜へ伸びる長い突起は残しており、これが皮質の上の層へ向かうニューロンの主要な足場になります。分裂の直前に細胞体が突起に沿ってジャンプするように移動する「分裂期細胞体転座(MST)」という独特の動きも見せます。
oRGが驚くべきなのは、脳室から遠く離れていて脳脊髄液からの栄養に頼れないにもかかわらず、自分で「幹細胞の居場所(ニッチ)」をつくり出す点です。oRGはテネイシンC・PTPRZ1・ITGB5・LIFRといった、成長シグナルを高める一連の遺伝子を強く働かせ、白血病阻止因子受容体(LIFR)とSTAT3という経路を自ら活性化して、爆発的な分裂を続けます[7]。このヒトに特徴的な「自前でニッチを維持する能力」の獲得こそが、高度な知性を支える大きく複雑な大脳皮質を生んだ最大の要因と考えられています。
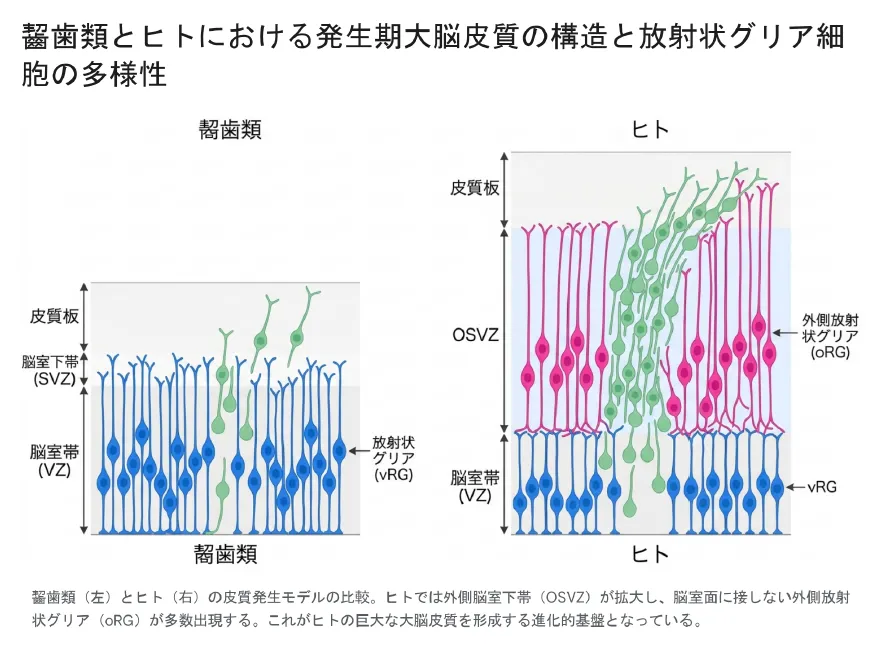
齧歯類(左)とヒト(右)の皮質発生の比較。ヒトでは外側脳室下帯(OSVZ)が拡大し、脳室面に接しない外側放射状グリア(oRG)が多数出現する。これがヒトの巨大な大脳皮質を形づくる進化的な基盤となっている。
なお、マウスにもわずかに基底放射状グリア様の細胞は存在しますが、正常な状態ではヒトのoRGのように何度も対称分裂を繰り返す力は持ちません。ヒトの皮質拡大に関わる分子としては、ヒトに特有の遺伝子であるARHGAP11Bが、基底側の前駆細胞を増幅させる因子として広く知られています。ARHGAP11Bは進化の過程でヒト系統に生じた遺伝子で、oRGを含む前駆細胞のプールを押し広げる働きがあると報告されています。こうしたヒト固有のしくみの積み重ねが、脳の巨大化を可能にしたと考えられています。

7. ニューロンを導く「足場」としての役割
放射状グリア細胞のもう一つの決定的な役割が、生み出したニューロンを大脳皮質の正しい層まで導く「足場(ガイドレール)」としての機能です。この「放射状遊走」は、静かなケーブルの上を細胞が滑るだけの単純な現象ではなく、細胞の骨組み・膜の動き・周囲のマトリックスが密に連携する、動的で精巧なプロセスです[10]。
ニューロンはまず脳室面から離れ(層間剥離)、いったん複数の突起を出す「多極性」の姿になった後、進行方向に1本の力強い突起を伸ばす「双極性」へと形を変えます。そして移動そのものは、細胞のかじ取り役である中心体が前へ引き上げられ、続いて核が中心体に向かって引き寄せられる、という段階的な牽引で進みます。この核の移動は、微小管という細いレールと、その上を動くモータータンパク質ダイニンの力に完全に依存しています。ここで働くLIS1とNDE1は、ダイニン複合体と協力して重い核を前へ引っぱる強力なエンジンを提供します[12]。ダイニンの重い部品をつくるDYNC1H1もこの遊走に関わります。
移動するニューロンが足場から滑り落ちずに前進と再接着を繰り返すためには、細胞どうしをつなぐ分子が欠かせません。アストロタクチンはニューロンとグリアの接着を仲介する中心的な分子で、この働きを抗体で妨げると、ニューロンの移動速度は平均で約61%も低下し、細胞内の物流も止まってしまうことが確かめられています[9]。またインテグリンは、ニューロン側と放射状グリア側でそれぞれ異なる種類が働き、細胞外マトリックスや細胞間の相互作用を仲立ちします。これらを阻害すると神経の遊走は大きく妨げられます[10]。
さらに最先端の知見として、放射状グリアの働きは物質を分泌するだけでなく、細胞外マトリックスそのものを組み立てることにも及びます。放射状グリアが出すテネイシンCとヒアルロン酸は、ニューロンが出すニューロカンと結びついて強力な「三元複合体」をつくり、移動中のニューロンを包む保護的・誘導的な足場として、多極性から双極性への切り替えを後押しします。遺伝子操作でこの複合体を壊すと、ニューロンの成熟が妨げられ、遊走に深刻な遅れが生じることが二重ノックアウトマウスで証明されています[8]。
遊走を語るうえで忘れてはならないのがリーリン(RELN)というタンパク質です。リーリンは皮質の層が「内側から外側へ」順序よく積み上がる(インサイド・アウト)パターンを制御し、遊走の最終段階でニューロンを正しい位置に着地させる合図を出します。リーリン経路の異常は、小脳の低形成を伴う滑脳症の原因にもなり、放射状グリアの足場機能と深く結びついています。放射状グリアの突起は、ギリシャ神話でテーセウスを迷宮から導いた「アリアドネの糸」にたとえられます——発生期の複雑な皮質という迷宮を、ニューロンが迷わず抜けるための確かな道標なのです[10]。
8. 血管ネットワークとの対話
放射状グリア細胞の仕事は、細胞を生み出し導くことだけにとどまりません。巨大な脳が生き延びるには、酸素と栄養を運ぶ丈夫な血管網が不可欠です。近年の研究で、放射状グリア細胞は皮質の血管づくり(血管新生)を空間的・時間的に制御する「オーガナイザー(調整役)」でもあることがわかってきました。
具体的には、放射状グリアは血管の内皮細胞に対してWnt/β-カテニンシグナルを適切に抑え、できたばかりの未熟な血管が退縮してしまうのを防いで構造を安定させます[11]。同時に、インテグリンβ8を介してTGF-βシグナルを局所的に働かせ、血管の枝分かれや内皮細胞の動きを微調整します。逆に血管側からも放射状グリアへフィードバックが返り、幹細胞の自己複製や分化のタイミングに影響を与えます。この双方向の対話が、神経づくりと血管づくりという2つのプロセスをぴたりと同期させ、健康な脳の「インフラ」を組み上げているのです。
9. 放射状グリアの不具合が引き起こす皮質形成異常
🔍 関連記事:大脳皮質形成異常NGSパネル/滑脳症NGSパネル/小頭症NGSパネル
放射状グリア細胞は「増やす力」と「導く力」という2つの大きな責務を単独で背負っています。だからこそ、これらの機能に遺伝子の変化による欠陥が生じると、大脳皮質の構造が根本から崩れ、重い発達の遅れ・運動障害・難治性てんかんを伴う皮質形成異常が起こります。原因遺伝子の多くは大脳皮質形成異常NGSパネルや神経細胞遊走障害NGSパネルなどで調べられます。
💡 用語解説:新生突然変異(de novo変異)とハプロ不全
新生突然変異(de novo変異)とは、両親のどちらにも無いのに、お子さんで初めて生じた遺伝子の変化のことです。皮質形成異常の多くはこのタイプで、家族歴がないことも珍しくありません。またハプロ不全とは、2つある遺伝子コピーの片方が働かなくなり、残り1つでは量が足りずに症状が出る状態を指します。滑脳症の代表的な原因の一つがこのしくみです。
① 小頭症:幹細胞プールの早すぎる枯渇
小頭症は、脳の発達が止まって頭囲が極端に小さくなる疾患です。中心となる病態は「放射状グリア(幹細胞)プールの早すぎる枯渇」で、とくに中心体や紡錘体の向きに関わる遺伝子の変異が致命的になります。ASPMやWDR62に変異が生じると分裂の紡錘体が異常な角度に傾き、前述したPar3の均等な分配のバランスが崩れます。その結果、幹細胞が自己複製する前に早まってニューロンへ分化してしまい、皮質ニューロンの総数が激減します[12]。WDR62は分裂期に紡錘体の極へ移動し、PLK1というキナーゼを中心体へ呼び込んで紡錘体の角度を安定させる役割が示されています[13]。またCDK6はoRGの細胞周期の進行に必須で、その異常も小頭症につながります。極端に重い型は微小滑脳症と呼ばれ、小頭症は小頭症NGSパネルで原因を調べます。
② 古典的滑脳症:ニューロンが登れなくなる
滑脳症は、脳のシワが形成されず表面が平らになる異常です。古典的滑脳症(1型)では、足場である放射状グリア自体は比較的保たれているのに、そこを登る「ニューロンの遊走エンジン」が壊れることで起こります。原因の中心は、微小管を安定させるDCXや、ダイニンを動かすLIS1の変異です[12]。LIS1が半分しか働かないハプロ不全になると、ニューロンは核を前へ引く力を失い、皮質表面まで登れずに白質などの異常な位置にとどまってしまいます。微小管の材料そのものであるTUBA1Aや、ARXの変異によるX連鎖滑脳症も知られます。LIS1を含む17p13.3領域の欠失はミラー・ディカー症候群を引き起こします。診断は滑脳症NGSパネルで行われます。
③ 敷石状滑脳症:足場が基底膜からはがれる
💡 用語解説:糖鎖修飾とグリア限界膜
糖鎖修飾とは、タンパク質に糖の鎖を取りつけて機能を整える「仕上げ加工」のことです。脳の最外層には、放射状グリアの末端足が付着する基底膜があり、ここが「ここで止まれ」という絶対的なバリア(グリア限界膜)になります。この付着を担うのが、糖鎖で仕上げられたα-ジストログリカンです。
敷石状滑脳症(2型)は、ニューロンが途中で止まるのではなく、逆に行き過ぎて(過剰遊走)脳表面に敷石状の凹凸をつくる病気です。α-ジストログリカンの糖鎖をつくるPOMT1・POMT2・FKRP・FKTN・POMGNT1などに変異が起きると糖鎖が欠け、放射状グリアの末端足が基底膜からはがれてバリアが崩壊します。すると停止信号を受け取れないニューロンがクモ膜下腔へあふれ出し、皮質を壊しながら異所性の細胞塊をつくります[14]。この病態はウォーカー・ワールブルグ症候群・筋・眼・脳病(MEB)・福山型先天性筋ジストロフィーという連続したスペクトラムを形づくり、ウォーカー・ワールブルグ症候群NGSパネルやジストログリカン関連先天性筋ジストロフィーNGSパネルで調べられます。
④ 脳室周囲結節性異所性灰白質(PVNH):足場そのものの崩壊
脳室周囲結節性異所性灰白質(PVNH)は、皮質へ向かうはずのニューロンの塊が移動を始められず、脳室の壁に沿って結節状にとどまってしまう疾患で、多くの患者さんがてんかんを発症します。最も一般的な原因は、X染色体上のFLNA(フィラミンA)の機能喪失変異です。フィラミンAはアクチンの骨組みを架橋して細胞の形と接着を保つタンパク質で、これが欠けると放射状グリアの極性が乱れ、脳室面の整然とした足場構造そのものが崩壊します[15]。足場が崩れると新生ニューロンはガイドレールを見つけられず、脳室周囲にたまって異所性の灰白質結節をつくります。より稀な常染色体潜性の型として、小胞輸送に関わるARFGEF2の変異による両側性PVNHも報告されています。
10. 成体脳・再生医療と、遺伝診療とのつながり
かつて放射状グリア細胞は「胎児期だけの一時的な存在で、生まれた後は消えてしまう」と考えられていました。しかし幹細胞生物学の進展により、成体の脳の限られた場所(脳室下帯や海馬歯状回)に残る「成体神経幹細胞」が、実は胎生期の放射状グリア細胞の性質を色濃く受け継いだ細胞であることがわかっています[2]。さらに、無脊椎動物のナマコなどでは、中枢神経が切断されると放射状グリア様細胞が活発に増殖して再生を主導することも確認されており、この細胞が動物界を通じて「組織の維持と再生」の核として保存されてきたことがうかがえます[2]。こうした知見は、脳オルガノイドを用いた病態モデルや、内在する幹細胞の力を引き出す次世代の再生医療への足がかりとして期待されています。
臨床の視点でみると、放射状グリア細胞は「用語」でありながら、遺伝診療の入口に直結しています。ここまで見てきた小頭症・滑脳症・敷石状滑脳症・PVNHは、いずれも胎児期の脳画像(超音波やMRI)で疑われることがあり、その先で原因遺伝子を調べる分子診断が診断確定と遺伝カウンセリングの土台になります。出生前の段階では羊水検査・絨毛検査による確定検査とターゲット遺伝子解析が、出生後には大脳皮質形成異常NGSパネルなどが用いられます。関連する多小脳回については多小脳回NGSパネルもあります。
これらの多くは前述の新生突然変異(de novo変異)で生じますが、X連鎖(DCX・FLNA・ARXなど)や常染色体潜性(多くの糖鎖修飾遺伝子)など遺伝形式はさまざまで、次のお子さんへの再発率も原因によって変わります。だからこそ、遺伝子の結果を家族の状況に即して読み解く遺伝カウンセリングが重要になります。ミネルバクリニックでは、こうした遺伝カウンセリングを臨床遺伝専門医が担当しています。
よくある質問(FAQ)
🏥 皮質形成異常・遺伝子診断のご相談
小頭症・滑脳症・敷石状滑脳症・PVNHなど
放射状グリアに関わる皮質形成異常の遺伝子検査・遺伝カウンセリングは
臨床遺伝専門医が在籍するミネルバクリニックにご相談ください。
参考文献
- [1] Radial glia – from boring cables to stem cell stars. Development. [Development]
- [2] Radial glia and radial glia-like cells: Their role in neurogenesis and regeneration. PMC. [PMC9708897]
- [3] Single Cell Analysis Reveals Transcriptional Heterogeneity of Neural Progenitors in the Human Cortex. PMC. [PMC5568903]
- [4] Molecular components and polarity of radial glial cells during cerebral cortex development. PMC. [PMC11105283]
- [5] Mammalian Par3 regulates progenitor cell asymmetric division via Notch signaling in the developing neocortex. PMC. [PMC2736606]
- [6] Hansen DV, Kriegstein AR, et al. Neurogenic radial glia in the outer subventricular zone of human neocortex. Nature. [Nature/NIG]
- [7] Molecular Identity of Human Outer Radial Glia During Cortical Development. PMC. [PMC4583716]
- [8] Assembly of neuron- and radial glial-cell-derived extracellular matrix molecules promotes radial migration of developing cortical neurons. eLife. [eLife 92342]
- [9] Astrotactin provides a receptor system for CNS neuronal migration. PubMed. [PubMed 1821847]
- [10] Ariadne’s Thread in the Developing Cerebral Cortex: Mechanisms Enabling the Guiding Role of the Radial Glia Basal Process during Neuron Migration. PMC. [PMC7822038]
- [11] Radial Glial Neural Progenitors Regulate Nascent Brain Vascular Network Stabilization Via Inhibition of Wnt Signaling. PLOS Biology. [PLOS Biology]
- [12] Genetics and biology of microcephaly and lissencephaly. PMC. [PMC3565221]
- [13] PLK1-mediated phosphorylation of WDR62/MCPH2 ensures proper mitotic spindle orientation. Human Molecular Genetics (Oxford). [Oxford HMG]
- [14] Cobblestone lissencephaly: neuropathological subtypes and correlations with genes of dystroglycanopathies. PubMed. [PubMed 22323514]
- [15] A glial origin for periventricular nodular heterotopia caused by impaired expression of Filamin-A. PubMed. [PubMed 22076441]






