目次
ヒトの卵子と精子が受精して着床するまでの過程をご紹介します。卵巣周期28日の14日目は排卵期で、その時期に数億の精子の中からたった一つと卵管膨大部で受精した胚子は7日かけて胚盤胞まで成長して子宮粘膜の細胞との接着分子どうしの相互作用により付着し、着床が成立します。
受精(fertilization)
受精とは男・女両生殖細胞の融合過程をさします。
受精は卵管膨大部でおこります。文字通り、卵管で一番幅広の部分で、卵巣にちかい位置にあります。
精子は女性の生殖管の中で数日間生存できます。
膣内の精子の1%だけが子宮頭部に入り、そこで長時間生存できます。子宮頸部から卵管までの精子の移動は、子宮と卵管の筋収縮によるところが大きく、精子自身の運動力の寄与はあまりありません。子宮頸部から卵管までの精子の移動時間は30分から6日間と様々です。卵管峡部に到達すると精子の動きは鈍くなり、移動が停止します。
排卵時期に精子はふたたび運動するようになるのですが、卵を取り囲む卵丘細胞が産生するケモカインによるものと考えられています。卵管峡部は受精が起こる卵管膨大部へと精子は泳いでいきます。
精子は、驚くことに女性生殖器へ到達した直後は卵子を受精させる能力を持ちません。
精子が受精できるようになるには、(1)受精能獲得、および(2)先体反応というステップが必要です。
受精能獲得(capacitation)
女性生殖路内で、精子を最良の状態にする調整期間で、ヒトでは約7時間ありますので、卵管膨大部に早く到達しても受精能獲得がまだなので、このような精子は卵子を受精させることができないため全く有利ではありません。巷ではよく、億単位の精子の中から力強く我先にと卵子に到達したものが勝ち、みたいな伝えられ方をしますが、誤りです。
受精能獲得の大部分は卵管内で精子と卵管粘膜上皮との相互作用で行われます。この期間中に、精子の先体域を担っている形質膜から糖蛋白質の被覆と精漿タンパクが除去されます。受精能を獲得した精子のみが放線冠を通過できます。
先体反応(acrosomereaction)
精子が透明帯に結合したのちに透明帯蛋白に誘導されて起こります。
この反応の頂点でアクロシンやトリプシン様物質を含む透明帯通過に必要な酵素類が放出されます。
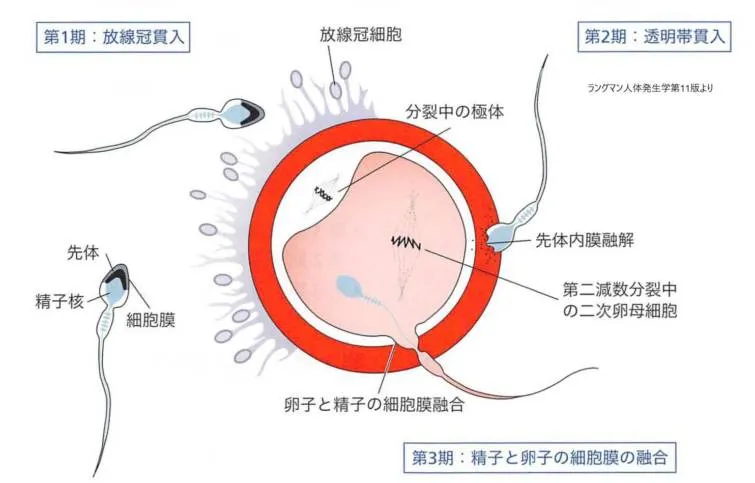
卵子貫通の3期を示す模式図です。
第1期:精子が放線冠関門を突破
第2期:1個またはそれ以上の精子が透明帯へ貫入
第3期:1個の精子が卵細胞膜に貫入。その際、精子自身の細胞膜は失う。
受精のステップ
受精には3期があります。
・第1期:放線冠貫入
・第2期:透明帯貫入
・第3期:卵子と精子の細胞膜の融合
第1期:放線冠貫入
女性の生殖路に送り込まれる精子は2億~3億個なのですが、膣から子宮に到達する時点で99%振るい落とされ、さらに受精の場である卵管膨大部に到達するのはわずか300~500個と言われています。これらの精子のうち受精に必要なのはl個だけで、その他は、受精する精子が卵子を保護する関門を通過するのを助けるために必要なのだと考えられています。
受精能を獲得した精子は、放線冠細胞の間を自由に通り抜けることが可能です。
第2期:透明帯貫入
透明帯は卵子を取り囲む糖蛋白質の殻で、精子の結合を促進、維持し、 先体反応を引き起こします。結合と先体反応は透明帯のタンパクであるZP3を介して起こりますが、ヒトでは生命倫理的な問題で研究に使える材料が乏しいためあまり解明が進んでいません。
先体の酵素であるアクロシンが放出されると、精子は透明帯を通過でき、卵子の細胞膜と接触出来るようになります。
透明帯の透過性は、精子の頭部が卵子の表面に接触すると変化し、卵子の細胞膜の直下にあった表層顆粒からリソソーム酵素が放出されます。この酵素は透明裕の性質を変える透明帯反応zona reactionを引き起こし、それ以降の精子の通過を妨害し、また透明帯表面に残っている精子に対する種特異性受容体部位を不活性化します。透明帯の中に他の精子が埋まっていることもありますが、卵子そのものに貫入できるのはl個の精子のみとなります。
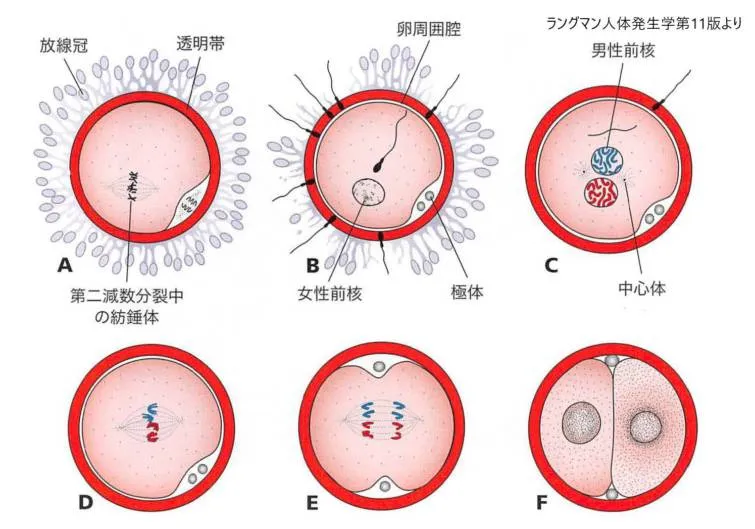
第3期:卵子と精子の細胞膜の融合
精子と卵子の初期の接着は、部分的には卵子表面のインテグリン(受容体)と精子表面のリガンドとの相互作用によっておこります。
接着に続いて、精子と卵子の細胞膜が融合します。
先体帽を覆っている細胞膜は先体反応中に消失するため、事実上の融合は卵細胞膜と精子頭部の後方部を覆う膜との間で行われます。ヒトでは、精子の頭部と尾部はいずれも卵子の細胞質内へ入るのですが、精子の細胞膜は卵子の表面に残されます。
精子が卵子に侵入し終わるとすぐ、卵子側の反応が起こります。
1. 表層反応および透明帯反応
リソソーム酵素を含む卵子表層頼粒の放出の結果以下の変化が起こります。
・卵細胞膜を他の精子が貫通できなくなる
・透明帯は精子の結合と貫通を阻害するように構造と組成が変化
こうして複数の精子が卵子に入り込む多精子受精が防止されます。
多精子受精を防止しなければ、通常2倍体のところ、3倍体以上の受精胚となり、発生できなくなるため、これを防止することは必須です。
2 . 第二減数分裂の再開
精子侵入直後に 卵子はその第二減数分裂を終了します。
2個の娘細胞のうち1つはほとんど細胞質を受け継がず、二次極体とよばれます。最終的に卵子となるのは1つだけです。その染色体(22,X)は女性前核(femalepronucleus)とよばれる胞状核内に配列します。
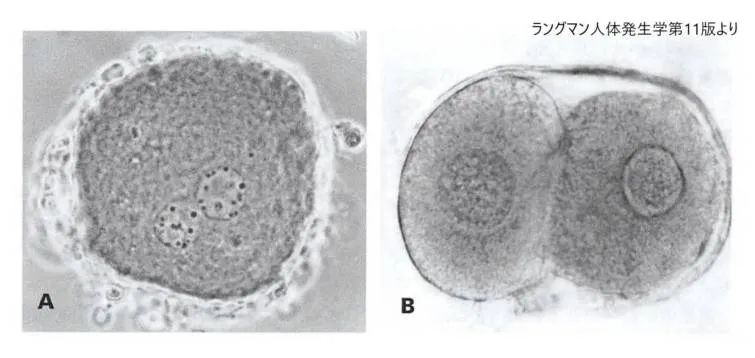
A 受精したヒト卵子の前核期の位相差顕微鏡像。男性および女性前核が確認できます。B ヒト2細胞期の接合
3.卵子の代謝活性化
活性化因子はおそらく精子が供給するものと想定されています。
活性化は、初期胚子発生に関連する最初の細胞的および分子的事象を含むものと考えられる。
その間に精子は前進して女性前核に接近し、核は膨張して男性前核を形成します。
精子の尾部は切り離され、男性前核と女性前核とは区別がつかず、最終的に両者は密着してその核膜を失います。
男性・女性前核(双方とも半数性)の発育中に各前絞はそのDNA量を倍加します。DNA合成の直後、染色体は正常のの有糸分裂に備えて、紡錘体上に並びます。母方、父方各23本の染色体(二重構造)が、動原体のところで縦に分裂し、姉妹染色分体が相対する極に向かつて移動し、こうして接合子の各細胞は、正常の染色体数と正怖のDNA量を与えられます。
姉妹染色分体が対側に移動する間に深い満が細胞の表面に生じ、しだいに細胞質を二分していきます。
分割(cleavage)
接合子が2細胞期に達すると、引き続き一連の有糸分裂が起きて細胞数が増加します。
これらの細胞は分割ごとに小さくなり、割球とよばれて8細胞期まではゆるやかに結合した塊となっています。
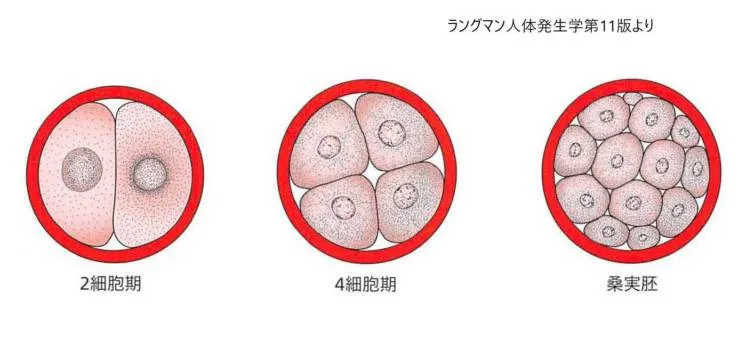
接合子が2細胞期から後期の桑実胚までの模式図。2細胞期には受精後約30時間、4細胞期は約40時間,12~16細胞期には約3日,後期桑実胚には約4日でそれぞれ到達します。この期間中、割球は透明帯で固まれていて、透明帯は受精後第4日の終わりには消失します。
しかし、第3分割に統いて、割球同士の結合が憎し、タイト結合でつながった細胞の密集した1つの球を形成します。
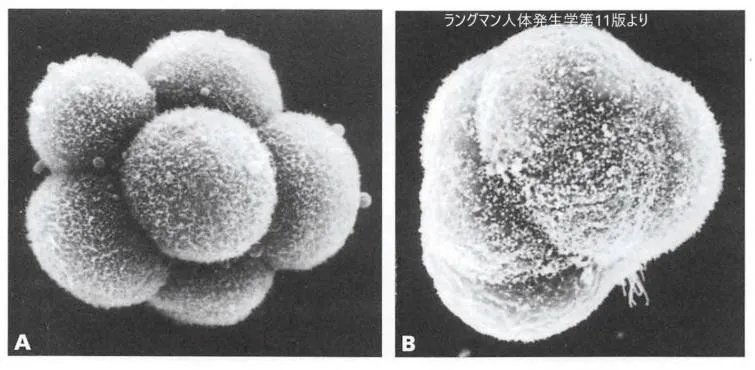
コンパクション前(A)コンパクション後(B)の8細胞期のマウス胚子走査電顕像です。コンパクション前には各
割球の輪郭は明瞭ですが、コンパクション後では細胞閣の接触が強固になり細胞の輪郭が不明瞭になっています。
この過程はコンパクション(compaction)とよばれ、ギャップ結合(隣り合う上皮細胞同士で水溶性の小さいイオンや分子を通過させる細胞間結合)を通じて盛んに情報交換している内部の細胞は、外部の細胞から分かれます。
受精後約3日でコンパクションを起こした胚子の細胞は再度分裂して16細胞の桑実胚(morula)となります。
桑実胚の内部にある細胞は内細胞塊(inner cell mass)をつくり、周囲を取り巻く細胞は外細胞塊を作ります。
内細胞塊からは胚子固有の組織が生じ、外細胞塊からはのちに胎盤形成をする栄養膜ができます。
胚盤胞の形成
桑実胚が子宮腔に入る頃に、液が透明帯を通り迎って内細胞塊の細胞の間隙に侵入し始めます。
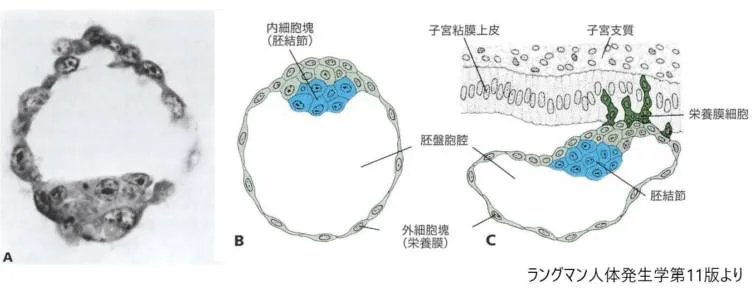
Aヒト胚盤胞(107細胞)の切片。内細胞塊と栄養膜細胞とが示されています。
B 子宮腔から得られた受精後約4.5日のヒトの腔盤胞の模式図。青色が内細胞塊(胚結節)、緑色が外細胞塊(栄養膜)を表す。
C 発生第6日の胚盤胞の模式図。胚盤胞の胚子極(動物極)に位置している栄養膜細胞が子宮粘膜に侵入し始めている様子が描かれています。ヒト胚盤胞は発生の第6日までに子宮粘膜に侵入し始めます。
ヒトの胚盤胞は100個程度の細胞でできています。青色の内細胞塊は赤ちゃんの身体のどんな部分にもなれる(成熟することを分化といいます)能力を持っています。
栄養膜は胎盤や羊膜などの胚外組織に分化していくことになります。
段々と細胞間隙は融合し、胚胞腔が形成され、胚盤胞となります。
内細胞脱の細胞は胚結節(embryoblast)とよばれ、一方の極に位置します。
外細胞塊の細胞は栄養膜と呼ばれ、扁平となり、胚盤胞の壁を形成します。
透明帯が消失し、着床を開始できるようになります。
ヒトでは、胚結節極を覆っている栄養膜上皮細胞は、発生のほぼ第6日に子宮粘膜上皮細胞間に侵入を始めます。
栄養膜上のL-セレクチンと子宮粘膜上皮上の炭水化物受容体が胚盤胞の子宮への初期接着に関与することが示唆されています。
セレクチンは白血球と血管内皮細胞間の機能分子で、E、L、Pセレクチンからなるファミリーを形成しています。そのうちLセレクチン(CD62L)は多くの白血球に発現し、Eセレクチン(CD62E)は炎症刺激により活性化した血管内皮細胞、Pセレクチン(CD62P)は活性化血小板や内皮細胞に発現すしています。
白血球と血管内皮細胞の接着メカニズムと同様のメカニズムが子宮上皮による胚盤胞の子宮腔からの補足の機構として提唱されています。
セレクチンによる捕捉に続いて、栄養膜細胞が発現するインテグリンと細胞外基質分子であるラミニンやフィブロネクチンとの相互作用により、接着と栄養膜の子宮粘膜内への侵入が促進されます。ラミニンとインテグリン受容体は付着を促進し、フィブロネクチンとインテグリン受容体は細胞の遊定を刺激します。これらの分子は、栄養膜の分化を調節するシグナル伝達経路で、この経路たちは相互に作用するため、着床は栄養膜と子宮内膜の相互作用がないとおこりません。
このようにしてヒトの接合子は、発生の第1週末までに桑実胚期と胚盤胞期を経過して、子宮粘膜に着床を開始します。
着床時の子宮
子宮の壁は3つの層からなります。
1.子宮内膜(endometrium):内壁を担う子宮粘膜
2.子宮筋層(myometrium):平滑筋の厚い層。
3.子宮外膜(perimetrium):外壁を覆う腹膜からなる層。
子宮粘膜の周期的な変化についてはこちらをクリックしてご覧ください。
着床時の子宮粘膜は分泌期にあたり、子宮腺と動脈はコイル状で、子宮組織は水分が多くなっていて、子宮内内膜には明瞭な3層が形成されています。表層の徹密層(compact layer)、 中間の海綿層(spongy layer)、基底層(basal layer)です。
通常、 ヒ卜の胚盤胞は子宮体の後壁または前壁の子宮内膜に着床します。
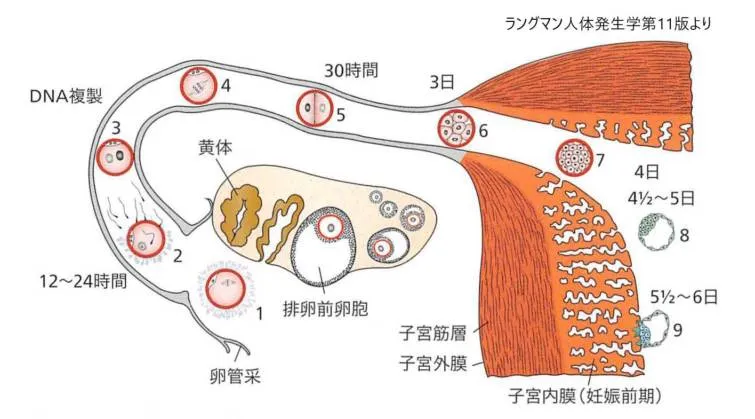
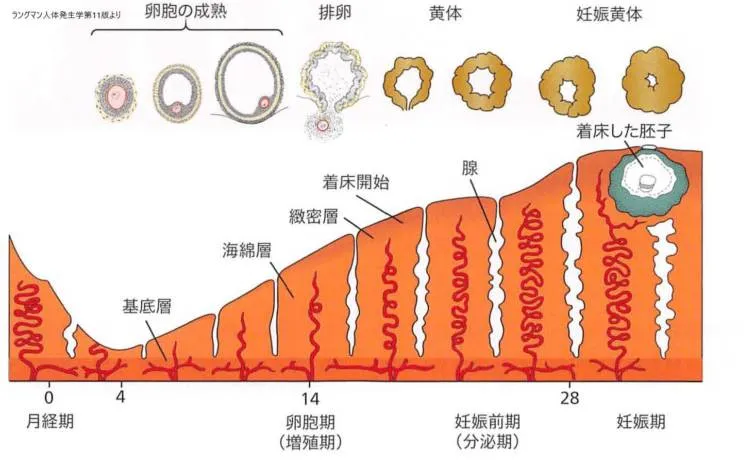
卵子が受精されないと、細動脈および洞様血管隙はしだいに血球がつまり、血液の広純な組織内流出がみられるようになります。月経期が始まると、表層の動脈から血液が流出し、支質と腺の小片が離脱します。これから3~4日のうちに、徹密層と海綿層が子宮粘膜から剥がれ、残るのは基底膜だけになります。