目次
- 1 1. クロモアナシンセシス(染色体再合成)とは
- 2 2. 3つのタイプ:破砕・錯綜・再合成はどう違う?
- 3 3. 分子メカニズム:複製の破綻とFoSTeS/MMBIR
- 4 4. 2026年の新発見:有糸分裂型MM-BIRの解明
- 5 5. 生殖細胞・初期胚での発生:父親由来と母親由来
- 6 6. 先天性疾患としての現れ方:ヤコブセン症候群など
- 7 7. がんとの関わり:ゲノムカオスと治療抵抗性
- 8 8. 進化生物学への波及:「有望な怪物」の復権
- 9 9. どうやって調べる?診断に用いる技術
- 10 10. 遺伝診療とのつながり:この用語はどこで役立つ?
- 11 よくある誤解
- 12 遺伝専門医からのメッセージ
- 13 よくある質問(FAQ)
- 14 参考文献
- 15 関連記事

「染色体がバラバラに壊れる」と聞くと、少しずつ傷が積み重なっていくイメージを持たれるかもしれません。しかしクロモアナシンセシス(染色体再合成)は、たった一度の細胞分裂という「一瞬」で染色体の一部が複雑に作り変えられる現象です。しかも壊れて散らばるのではなく、DNAをコピーする仕組み(複製)のエラーによって、同じ場所が二重・三重に増える点が特徴です。このページでは、その分子の仕組みから、2026年に解明された最新の発見、先天性疾患やがん、さらには進化との関わりまでを、遺伝専門医の視点でやさしく解説します。
Q. クロモアナシンセシスとは何ですか?まず結論だけ知りたいです
A. クロモアナシンセシス(染色体再合成)とは、DNAを複製する仕組みが一度の細胞分裂の中で破綻し、その結果として染色体の一部が複雑に作り変えられる現象です。物理的に砕けるのではなく、複製が別の場所へ「飛び移る」ことで、同じ領域が重複・三重複(コピー数の増加)を起こすのが最大の特徴です。先天性疾患やがんの原因になるほか、生物の進化を一気に進める仕組みとしても注目されています。
- ➤正体 → DNA複製のエラー(FoSTeS/MMBIR)で起こる、複製ベースの複雑な染色体再構成
- ➤仲間との違い → クロモスリプシス(破砕)・クロモプレキシー(錯綜)と並ぶ「クロモアナジェネシス」の一員
- ➤2026年の新発見 → 有糸分裂型MM-BIRという仕組みが解明され、暴走の全容が明らかに
- ➤臨床との関わり → 先天性疾患・発達遅滞・がんの成り立ちに関与し、精密な検査が診断の鍵
- ➤調べ方 → 染色体マイクロアレイ・全ゲノム解析・ロングリードなど新世代の技術が必要
1. クロモアナシンセシス(染色体再合成)とは
私たちの体をつくる細胞は、分裂のたびに約30億塩基対のDNAを正確にコピーしています。このコピー作業が一度の細胞分裂の中で大きく破綻し、その結果として染色体の一部が複雑に再構成されてしまう現象をクロモアナシンセシス(染色体再合成)と呼びます。語源はギリシャ語で「染色体(chromo)」と「再合成・再構成(anasynthesis)」を組み合わせたもので、名前のとおり、DNAを合成し直す過程で起こる異常です。
長い間、がんの進行や生物の進化は、小さな変異が何万年もかけて少しずつ積み重なって生じる、と考えられてきました。ところが次世代シーケンサーや高解像度の染色体マイクロアレイが普及すると、たった一度の破局的なイベントで、ゲノムが一気に大規模に組み替わる現象が次々と見つかりました[1]。クロモアナシンセシスは、その代表格のひとつです。
💡 用語解説:クロモアナジェネシス(染色体新生)
クロモアナジェネシス(chromoanagenesis)とは、「一度の破局的なイベントで起こる大規模な染色体再構成」の総称です。日本語では「染色体新生」「染色体再生」などと訳されます。この大きな傘のなかに、クロモスリプシス(破砕)・クロモプレキシー(錯綜)・クロモアナシンセシス(再合成)という3つのタイプが含まれます。名前がよく似ていますが、起こる仕組みも、できあがる染色体の姿も、それぞれ大きく異なります。より広い枠組みはクロモアナジェネシスの解説ページでも紹介しています。
クロモアナシンセシスがほかの2タイプと決定的に違うのは、染色体が物理的な衝撃で「砕ける」のではなく、DNAをコピーする複製という作業のエラーから生まれる点です。この違いは、できあがるゲノムの形に直接あらわれます。砕けて再結合するタイプでは欠失(DNAが失われること)が目立つのに対し、クロモアナシンセシスでは同じ領域が二重・三重にコピーされる局所的な重複・三重複(コピー数の増加)が特徴として残ります。この「増える」という指紋こそ、クロモアナシンセシスを見分ける手がかりになります。
クロモアナシンセシスという概念が整理されたのは、比較的最近のことです。2010年代の初めに、がんゲノムのなかで「少しずつ」ではなく「一度に」大規模な再構成が起こる現象が相次いで報告され、研究が一気に加速しました。その流れのなかで、複製の仕組みが暴走して重複を生むタイプを指す言葉として、クロモアナシンセシスが位置づけられました。それまで「原因不明の複雑な染色体異常」とひとくくりにされてきた多くのケースが、じつは一度きりの破局的なイベントの産物だった、と理解し直されつつあります。こうしたコピー数変異が一か所に密集して現れる姿は、従来の「変異は少しずつ積み重なる」という考え方だけでは、うまく説明できないものでした。
2. 3つのタイプ:破砕・錯綜・再合成はどう違う?
3つのタイプは、関わる染色体の本数、コピー数がどう変わるか、どのDNA修復経路を使うか、という点で明確に線引きできます。まずは全体像を表で整理します。
言葉の由来をたどると違いがよくわかります。クロモスリプシスは「粉砕(thripsis)」、クロモプレキシーは「編み込む・ねじれる(pleko)」、そしてクロモアナシンセシスは「再合成(anasynthesis)」です。クロモスリプシスが「割れたガラスを貼り合わせる」イメージだとすれば、クロモアナシンセシスは「コピー機が別ページに飛んで印刷し続け、同じ段落が何度も重複してしまう」イメージです。下の図は、この3タイプの構造的な違いを示したものです。
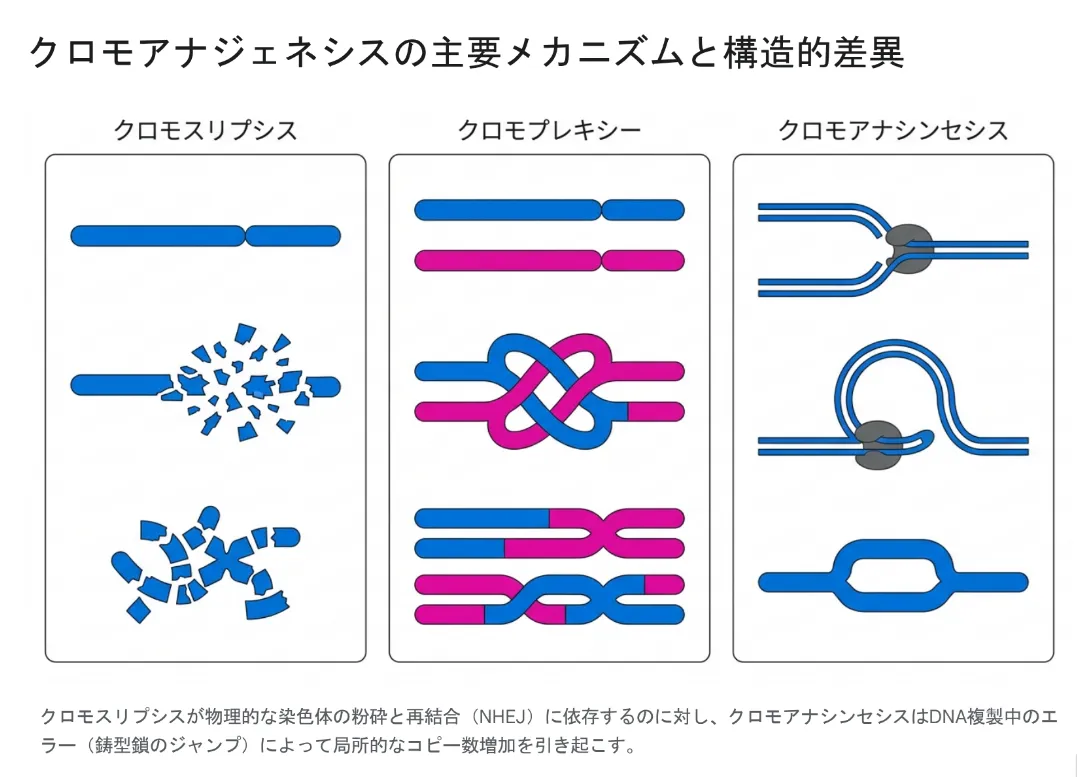
クロモスリプシスが物理的な粉砕と再結合(NHEJ)によって起こるのに対し、クロモアナシンセシスはDNA複製中のエラー(鋳型鎖のジャンプ)によって局所的なコピー数の増加を引き起こします。
3. 分子メカニズム:複製の破綻とFoSTeS/MMBIR
クロモアナシンセシスを理解する鍵は、細胞周期のS期(DNAをコピーする時期)に起こる複製の破綻です。活性酸素・化学物質・放射線といった外的ストレスや、ゲノム上の「コピーしづらい配列」(脆弱部位、強固なヘアピン構造、反復配列など)があると、DNAをコピーしていく装置、すなわち複製フォークが停止したり、崩壊したりします。通常なら相同組換えという正確な修復でリカバリーしますが、それが難しいと、細胞はエラーを起こしやすい代替経路へ切り替えざるを得なくなります[2]。
💡 用語解説:FoSTeS/MMBIR
FoSTeS(Fork Stalling and Template Switching:複製フォークの停止と鋳型鎖の乗り換え)と、それをより一般化したMMBIR(Microhomology-Mediated Break-Induced Replication:微小相同性を介した切断誘発複製)は、クロモアナシンセシスを説明する中心的なモデルです。止まってしまった複製が、元の場所に戻る代わりに、たまたま近くにある別の場所へ「飛び移って」コピーを続けてしまう——この乗り換えが繰り返されることで、ゲノムのあちこちの断片をつなぎ合わせた複雑なパッチワークが生まれます。詳しくはFoSTeS/MMBIRの解説ページもご覧ください。
停止した複製フォークでは、合成途中だったDNA鎖の端(3’末端)が露出します。正常なら元の鋳型に戻るのですが、クロモアナシンセシスでは、この露出した鎖が三次元的に近くにある別の複製フォークへ侵入し、そこを新しい鋳型としてコピーを続けてしまいます。重要なのは、飛び移る先が染色体上で隣り合っている必要はなく、核のなかで物理的に近ければよいという点です。しかもこの異常なコピーは不安定なため、乗り換えが何度も繰り返されます。こうして最終的にできあがる染色体は、遠く離れた領域の配列をつなぎ合わせた複雑なつぎはぎとなり、局所的な重複や三重複を抱え込むことになります。複製ストレスが、この一連の暴走の引き金になっているのです。
💡 用語解説:微小相同性(マイクロホモロジー)
露出したDNAの端が別の場所へ飛び移るとき、足がかりとして使われるのがわずか3〜5塩基対のごく短い一致配列(微小相同性)です。この短い一致が接合部に残っていることは、その再構成が「単純に切って貼った(NHEJ)」のではなく「複製の仕組みが暴走した(MMBIR)」ことを示す決定的な分子の指紋になります。ヒトのゲノムには反復配列がとても豊富で、この短い一致配列を無数に提供してしまうことが、乗り換えが起こりやすい背景にあると考えられています。
ではなぜ、細胞は正確な相同組換えではなく、エラーの多い複製依存の経路を選んでしまうのでしょうか。相同組換えという正確な修復には、コピーの手本となるよく似た配列(姉妹染色分体)と、十分な時間・修復酵素が必要です。ところが複製ストレスによって多数の複製フォークが同時に止まってしまうと、手本が間に合わず、細胞は「不正確でも先へ進む」MMBIRに頼らざるを得なくなります。いわば非常事態の応急処置として選ばれた経路が、結果的に大きな傷跡を残してしまうのです。さらにヒトのゲノムは、AluやLINEと呼ばれる反復配列がゲノム全体のおよそ半分を占めるほど豊富で、これらが3〜5塩基対の短い一致配列を至るところに提供します。乗り換え先の「足がかり」がゲノム中に無数に散らばっていることも、クロモアナシンセシスがヒトで起こりやすい一因だと考えられています。
4. 2026年の新発見:有糸分裂型MM-BIRの解明
🔍 関連ページ:MMEJ(微小相同性媒介末端結合)/サブテロメア/ロングリードシーケンシング
長らく詳細が不明だった「クロモアナシンセシスを実際に駆動している分子の正体」が、2026年3月に大きく前進しました。カーディフ大学のGreg Ngo博士とDuncan Baird教授らのチームが、一分子ロングリードシーケンシングを用いて、この破局的な再構成が起こる瞬間を高い解像度でとらえ、Nature Communications誌に報告したのです[4]。
この研究が明らかにしたのは、クロモアナシンセシスが「有糸分裂(細胞が実際に2つに分かれる段階)」で特異的に働くMM-BIRによって生じる、という点です。とくに、短くなったテロメアや、テロメアのすぐ内側(サブテロメア)で起きたDNA二本鎖切断のときに、この経路が作動しやすいことがわかりました。通常、細胞は分裂に入る前のチェックポイントで傷を直すか、直せなければ自ら死を選びます。しかし発がんの初期などでこのチェックポイントが働かなくなると、傷を抱えたまま分裂に突入し、分裂直前に強引に修復しようとする「非常用の安全装置」としてMM-BIRが暴走するのです。
さらに驚くべきことに、この経路はMMEJ(微小相同性媒介末端結合)とBIR(切断誘発複製)という、これまで別々に考えられていた2つの修復モードが緊密に連携して進むことが示されました。MMEJに関わるタンパク質群が、DNAポリメラーゼδ(Polδ)に依存したBIRを開始させ、それをPIF1・POLD3・PCNAといった因子が調節します。この乗り換えのしやすさゆえに、ときには逆方向反復配列に複製が「捕まって」しまい、たった一度の分裂で単一の領域が何十倍にも異常増幅する、想像を絶するゲノムの混乱が引き起こされます。MMEJとBIRが要素を共有していることは、同じ細胞のなかでクロモアナシンセシスとクロモスリプシスがしばしば同時に見つかる理由も、うまく説明してくれます。
この2026年の研究がもつ意味は小さくありません。従来の短く読む解析では、複雑に絡み合った領域は「読めない空白」として残ってしまい、何がどうつながっているのかを正確に描けませんでした。ロングリードで一本のDNAを端から端まで読み通せるようになってはじめて、乗り換えの回数や順序、そして接合部の指紋を一つずつ確かめられるようになったのです。仕組みが分子レベルで見えてきたことは、将来、この暴走を早期に見つけたり、抑えたりする手がかりを探すうえでの重要な出発点になります。

5. 生殖細胞・初期胚での発生:父親由来と母親由来
🔍 関連ページ:高齢父性(父親の加齢)/染色体不分離/減数分裂とは
クロモアナシンセシスはがん細胞で最初に注目されましたが、近年は正常なヒトの生殖細胞(精子・卵子)や初期胚でも起こることがわかってきました[3]。生殖細胞で起きた変化は次の世代のすべての細胞に受け継がれるため、影響は非常に大きくなります。興味深いことに、その成り立ちは父親由来か母親由来かで大きく異なります。
初期に報告された重複・三重複型の複雑な再構成は、父親由来(精子起源)が優勢でした。男性の精子のもとになる細胞(精原細胞)は、生涯にわたって分裂を続けます。50歳になるまでに1000回を超える分裂を重ねることもあり、そのぶん複製エラーが起こる確率が積み上がっていきます。さらに精原細胞は、傷があっても細胞死を避けてコピーをやり遂げる特殊な仕組みを持っており、これが結果的にFoSTeS/MMBIRの活性化につながると考えられています。父親の加齢とともに新生突然変異のリスクが高まることと、同じ背景を共有しています。
一方、女性の卵子のもとになる細胞は、胎児期にしか分裂せず、その回数は30回に満たないほどです。そのため「分裂回数の多さによる複製エラー」のリスクは本質的に低いのですが、代わりに染色体の分配異常(うまく振り分けられないこと)が主な要因になります。卵母細胞は何十年も休止状態に置かれるため、加齢とともに染色体を束ねるタンパク質が劣化し、分裂の際に染色体が取り残される「遅行染色体」が生じやすくなります。
💡 用語解説:微小核(マイクロニュークリアス)
分裂のときに取り残された染色体は、本体の核に入りきれず、細胞質のなかに小さな別の核をつくることがあります。これが微小核です。微小核の膜はとても壊れやすく、内部の環境も劣悪なため、そこに閉じ込められた染色体は未熟なまま凝縮されて物理的に砕けたり、激しい複製ストレスにさらされてFoSTeS/MMBIRを暴走させたりします。この微小核こそが、染色体の粉砕と異常な複製を起こす「巨大なインキュベーター(培養器)」の役割を担うことが、最新の生細胞イメージングで示されています。
この微小核の崩壊を起点に、一度の細胞周期で大規模な染色体再構成が一気に生成されます。なお、体外受精や顕微授精といった生殖補助医療(ART)の培養環境で生じる物理的・化学的なストレスが、こうした微小核形成やクロモアナシンセシスに影響しうるかどうかは、現在も活発に研究が続けられている段階です。断定できる結論はまだ得られていません。
なお、受精した直後の胚の細胞でクロモアナシンセシスが起こった場合は、その細胞から分かれた一部の細胞だけが変化を抱える「モザイク」の状態になります。体のどの組織に、どの割合で広がるかによって、症状の重さや現れ方が大きく変わるため、まったく同じ再構成でも一人ひとりで経過が異なります。この点が、生まれつきのクロモアナシンセシスの見通しを一律に語れない理由のひとつになっています。
6. 先天性疾患としての現れ方:ヤコブセン症候群など
生殖細胞や初期発生の段階でクロモアナシンセシスが起こると、その変化があまりに大きいため、多くの場合は細胞死や流産に至り、赤ちゃんとして生まれてくることができません。再構成されたゲノムが、生存に必須の遺伝子を致命的に壊しておらず、機能的に「ぎりぎり許容できる」まれなケースだけが個体として成立し、さまざまな先天性疾患や発育の違いとして現れます。症状は影響を受けたゲノム領域によって大きく異なり、特徴的な顔立ち、自閉スペクトラム症、知的障害、発達の遅れから、比較的軽いものまで幅広く見られます。
ここで大切なのは、生まれつきのクロモアナシンセシスが「まれにしか生き残れない」という点です。大規模な再構成の大半は、発生のごく初期に細胞死や流産という形で自然に淘汰されます。そのため、実際に診断にたどり着く症例は、厳しいふるいをくぐり抜けたごく一部にすぎません。この「氷山の一角」という性質こそが、これまでこの現象が長く見過ごされてきた大きな理由のひとつでもあります。次世代の解析技術が普及したことで、ようやく水面下に隠れていた全体像が見えはじめたところです。
先天性のクロモアナシンセシスが特定の症候群を直接引き起こしたと証明された象徴的な例が、ヤコブセン症候群です[6]。この病気は通常、11番染色体の長腕(11q)の末端が失われ、170から340以上の遺伝子が欠けることで起こります。ところが、ある患者さんの詳しい解析で、たった一つのストレスを受けた細胞で生じたクロモアナシンセシスが原因だったことが判明しました。
この患者さんの11qでは、片方のコピーに6か所の重複と5か所の欠失が集中して起きていました。合計すると約4.894メガベースが重複した一方で、別の領域では約6.8メガベースが失われ、差し引き約2メガベースの正味のDNA喪失が生じていたのです。この過程で、ヤコブセン領域にあった18個の遺伝子が欠失し、そのなかに知的障害と関連する中核遺伝子KIRREL3が含まれていました。接合部には明確な微小相同性の指紋があり、この再構成が段階的な積み重ねではなく、複製ベースの「オール・アット・ワンス(一度に全部)」のイベントだったことが確定しました。
補足:ヤコブセン症候群の従来型は、単純な11q末端欠失が原因です。上記はあくまで「クロモアナシンセシスによって同じ症候群が生じた特殊な一例」であり、すべてのヤコブセン症候群がこの仕組みで起こるわけではありません。関連情報は11q23欠失症候群のページもご参照ください。
クロモアナシンセシスは、ヒトゲノムのさまざまな領域で報告され続けています。なかには表現型が正常な個体で見つかる例もあります。てんかんや運動失調を示す小児で21番染色体にクロモアナシンセシス領域が見つかったものの、症状のない母親や祖父も同じ不均衡な染色体を持っており、最終的に小児の症状は再構成領域内のSYNGAP1遺伝子の病的バリアントによると考えられました[9]。同じ構造変化でも、どの遺伝子がどう影響を受けるかで結果が大きく変わることを示す例です。
2026年には、長らく細胞遺伝学的な診断が難しかった構造変異のひとつ「過剰マーカー染色体(sSMC)」に、最新のロングリード・ゲノムシーケンシングが応用されました。臨床で見つかった10例のsSMCを解析した結果、そのうち9例でセントロメアの不安定性とクロモアナシンセシスが関与していることが示され、複雑なタイプではトリソミーレスキュー(余分な染色体を細胞が処理しようとする過程)の破綻が主要な形成メカニズムとして実験的に裏づけられました[5]。原因不明とされてきた染色体異常症候群の一部が、発生初期の複製ベースの破局的イベントに由来する可能性を強く示唆しています。
7. がんとの関わり:ゲノムカオスと治療抵抗性
🔍 関連ページ:がん遺伝子(オンコジーン)/がん抑制遺伝子/遺伝子増幅
生まれつきの影響とは対照的に、体の細胞で後天的に起こるクロモアナシンセシスは、がんの発生と悪性化に決定的な役割を果たします。がん細胞は、点突然変異を長い時間かけてためこむだけでなく、一度の細胞周期でゲノムの設計図を一気に書き換える能力を持ちます。この「ゲノムカオス」の局面は、がん細胞に治療への即座の抵抗性と、急速な悪性化の機会を与えてしまいます。
象徴的なのが、まれな腎平滑筋肉腫の症例解析です。腫瘍ゲノム内に3つの明確なクロモアナシンセシス領域が見つかり、MDM2やCDK4といった細胞周期を暴走させるがん遺伝子の強烈なコピー数増加が引き起こされていました。重要なのは、この腫瘍ではTP53・RB1などの主要ながん抑制遺伝子には点突然変異が一切見られなかったことです。クロモアナシンセシスによる染色体不安定性そのものが、発がんの直接の引き金になったと推測されています[7]。小児・青少年に多い骨肉腫でも、クロモアナシンセシスやクロモスリプシスが必須の腫瘍抑制遺伝子を一瞬で欠失させ、がん遺伝子を異常増幅させることで、治療抵抗性の強いサブクローンの出現を後押しすることが指摘されています[8]。
クロモアナシンセシスと他の複雑なイベントが重なると、コピー数変異の影響を受ける遺伝子の数が跳ね上がります。下のグラフは、その激増ぶりを示したものです。
複雑なゲノム再構成が生むコピー数変異遺伝子の激増
各イベント群における、コピー数変異(CNA)を持つ平均遺伝子数の比較
再構成なし
クロモスリプシスのみ
その他の複雑なイベント
複合したイベント
複数の複雑なイベントが重なった群では、コピー数変異の影響を受ける遺伝子数が、再構成のない群と比べておよそ2倍以上に急増します。
生み出されるゲノムの変化は文字どおり「カオス的」ですが、生き残る細胞が完全にランダムに選ばれるわけではありません。生存を支えられるのは、細胞の代謝や生存に直結する必須機能をかろうじて維持できた再構成だけです。膨大な細胞集団のなかでごくまれに、致命的な欠損を免れつつ増殖能力を得た「機能的に許容できる」ゲノムが誕生し、それが治療への強い適応力を持って爆発的に進化していきます。血液のがん(骨髄異形成症候群や急性骨髄性白血病など)でも、染色体マイクロアレイ検査とTP53変異の検索を組み合わせ、こうした高リスクな病態をすばやく確実にとらえることが、今の臨床の課題となっています。
がん治療の観点から見過ごせないのは、この一度きりの大再編が、治療への抵抗性を一瞬で獲得する近道になりうる点です。増幅されたがん遺伝子が、染色体から飛び出した小さな環状のDNA(染色体外DNA)として細胞内で増えることもあり、これが薬剤への耐性を生む温床になると知られています。さらに、こうした複雑な変化がゲノムに残した痕跡のパターンは、変異シグネチャーとして解析され、どの仕組みが働いた結果なのかを推定する手がかりになります。破局的なイベントの「筆跡」を読み取る試みは、がんの成り立ちを理解するうえで欠かせない視点になりつつあります。
8. 進化生物学への波及:「有望な怪物」の復権
クロモアナシンセシスの発見は、医学やがん研究の枠を超えて、進化生物学の根本的な考え方にも波及しています[10]。ダーウィン以来の伝統的な進化論は、小さな変異が数百万年かけて少しずつ積み重なることで新しい種が生まれる、と説明してきました。しかしクロモアナシンセシスは、たった一回の細胞分裂という「まばたきするような時間」でゲノム全体が構造的に作り変えられる現象であり、この漸進的なモデルとは真っ向から対立します。
この概念は、1940年代に遺伝学者リチャード・ゴールドシュミットが提唱し、長らく異端とされてきた説に、分子レベルの裏づけを与えるものです。彼は、種のあいだの大きな違いは微小な変異の蓄積では説明できず、初期発生を劇的に変える大規模な変異が必要だと主張し、そこから生まれる新しい可能性を持つ個体を「Hopeful Monsters(有望な怪物)」と呼びました。また、進化が長い安定期と短い爆発的変化の時期を繰り返すとする「断続平衡説」にも、現代的な証拠を与えています。実際、ヒトだけでなく、哺乳類全般や真菌、植物など多様な生物の進化の痕跡に、クロモアナジェネシス由来の大規模再構成が確認されつつあります。ただし、これらは基礎研究として明らかになりつつある知見であり、進化の全体像における位置づけは今も議論が続いています。
クロモアナシンセシスは、病気の原因という「影」の側面だけでなく、生命が一気に新しい形を生み出しうるという「光」の側面も併せ持っています。破壊と創造が同じ一つの仕組みの表と裏になっているという事実は、ゲノムというものへの見方そのものを、私たちに静かに問い直させてくれます。
9. どうやって調べる?診断に用いる技術
🔍 関連ページ:染色体マイクロアレイ(CMA)/オプティカルゲノムマッピング/核型(カリオタイプ)
クロモアナシンセシスのような超複雑な構造変異は、従来の細胞遺伝学的検査だけでは十分にとらえられません。前述のヤコブセン症候群の例でも、Gバンド法による核型分析やFISH法では、単純な末端欠失しか見えず、内部の複雑な再構成は把握できませんでした。ゲノムカオスを正確に読み解くには、次のような新世代のアプローチが欠かせません。
クロモアナシンセシスの多くは、全体として遺伝物質の量が釣り合わない不均衡型の再構成として現れます。どの領域が、どれだけ増え、どれだけ失われているのか——この不均衡をどれほど細かく正確に地図化できるかが、検査技術の性能を分ける決め手になります。
🔬 染色体マイクロアレイ(CMA)
コピー数の増減(重複・欠失)の位置と大きさを正確にマッピングします。クロモアナシンセシス最大の特徴といえる「局所的なコピー数増加のクラスター化」をとらえる、臨床の第一選択ツールです。
🧬 全ゲノム解析(WGS)
80倍以上の高精度で読むことで、CMAでは見えない均衡型の転座や逆位も検出できます。さらに接合部の微小相同性の指紋を1塩基単位で特定し、原因がNHEJかFoSTeS/MMBIRかを見分けられます。
🧪 ロングリード/OGM
長いDNAを一気に読むロングリードや、オプティカルゲノムマッピングは、複雑に絡み合った領域や反復配列の再構成を端から端まで解明します。2026年のsSMC研究で威力を発揮しました。
これらの技術は、単に「異常がある」ことを示すだけでなく、その異常がどんな仕組みで生じたのかまで教えてくれます。仕組みがわかれば、再発のリスクをより正確に説明でき、ご家族への情報提供の質も高まります。当院では染色体マイクロアレイ検査や全ゲノム解析、また染色体不安定症候群の検査についてもご案内しています。
10. 遺伝診療とのつながり:この用語はどこで役立つ?
クロモアナシンセシスは基礎研究の言葉に見えますが、実際の遺伝診療の3つの場面と深くつながっています。ひとつめは出生前診断・着床前診断で複雑なコピー数の変化が見つかったときの解釈です。重複・三重複がクラスターになっている所見は、この現象を念頭に置くことで意味づけが変わってきます。ふたつめは原因不明の発達の遅れや多発奇形に対する検査選択で、CMAや全ゲノム、ロングリードといった技術が診断の突破口になります。みっつめは遺伝カウンセリングでの再発リスクの説明で、新生突然変異なのか、生殖細胞のモザイクが関わるのかによって、次のお子さんへの見通しが変わります。
こうした説明や検査の選択、結果の解釈は、臨床遺伝専門医が担う領域です。染色体の複雑な再構成という難しいテーマを、ご本人やご家族にわかりやすい言葉でお伝えし、次の一歩を一緒に考えていくことを大切にしています。
よくある誤解
誤解①「染色体が物理的に砕ける現象でしょう?」
砕けて再結合するのはクロモスリプシスです。クロモアナシンセシスは、DNAをコピーする過程のエラー(複製ベース)で起こり、同じ領域が増える(重複・三重複)のが特徴です。似ていますが仕組みは別物です。
誤解②「がんだけで起こる特殊な現象では?」
最初はがんで注目されましたが、正常な精子・卵子や初期胚でも起こることがわかっています。生まれつきの染色体異常や発達の違いの原因になる場合もあります。
誤解③「見つかったら必ず重い病気になる?」
大規模な再構成の多くは細胞死や流産に至ります。生存する例の症状は影響領域により幅広く、症状のないまま同じ染色体を持つご家族が見つかることもあります。
誤解④「普通の染色体検査で分かる?」
Gバンド法やFISHでは内部の複雑さを十分にとらえられません。CMA・全ゲノム解析・ロングリードなど新世代の技術ではじめて全体像が見えてきます。
遺伝専門医からのメッセージ
よくある質問(FAQ)
🏥 染色体・遺伝子のご相談
複雑な染色体の所見や、原因のわからない発達の違いについて
遺伝子検査・遺伝カウンセリングをご検討の方は
臨床遺伝専門医が在籍するミネルバクリニックにご相談いただけます。
参考文献
- [1] Pellestor F. Chromoanagenesis: the mechanisms of a genomic chaos. Semin Cell Dev Biol. [PubMed 33608210]
- [2] Liu P, et al. The DNA replication FoSTeS/MMBIR mechanism can generate genomic, genic and exonic complex rearrangements in humans. Nat Genet. [PMC4461229]
- [3] Chromoanasynthesis: another way for the formation of complex chromosomal abnormalities in human reproduction. Hum Reprod. [Oxford Academic]
- [4] Ngo GHP, et al. Mitotic microhomology-mediated break-induced replication promotes chromoanasynthesis. Nat Commun. 2026;17:3375. [Nature Communications]
- [5] Bilgrav Saether K, et al. Centromeric instability and chromoanasynthesis observed in nine supernumerary marker chromosomes resolved with long-read genome sequencing. Genome Res. 2026;36(4):661-670. [Genome Research]
- [6] Chromoanasynthesis as a cause of Jacobsen syndrome. Am J Med Genet A. [PMC11007684]
- [7] Multiple Chromoanasynthesis in a Rare Case of Sporadic Renal Leiomyosarcoma. Front Oncol / PMC. [PMC7466669]
- [8] Chromoanagenesis in Osteosarcoma. Biomolecules. [MDPI Biomolecules]
- [9] The Iceberg under Water: Unexplored Complexity of Chromoanagenesis in Congenital Disorders. Genes / PMC. [PMC6451730]
- [10] Pellestor F, et al. Chromoanagenesis: a piece of the macroevolution scenario. Mol Cytogenet / PMC. [PMC6988253]






