目次

📍 クイックナビゲーション
DNAが真っ二つに切れる「二重鎖切断」は、細胞にとって最も危険な傷です。この傷を修復する経路は長らく2つと考えられてきましたが、近年、両者が働けない場面で細胞を救う「第三の経路」=MMEJ(マイクロホモロジー媒介性末端結合)の重要性が明らかになりました。MMEJはDNAポリメラーゼθ(POLQ)という特殊な酵素が主役で、少しの欠失や挿入を残す「誤りがち」な性質を持ちます。この記事では、MMEJの仕組みから、BRCA変異がんに対する新しい治療戦略、CRISPRゲノム編集への応用まで、臨床遺伝専門医がやさしく解説します。
Q. MMEJ(マイクロホモロジー媒介性末端結合)とは何ですか?まず結論だけ知りたいです
A. MMEJは、切れたDNAの両端にある「1〜25塩基ほどのごく短い共通配列(マイクロホモロジー)」を目印につなぎ合わせる、DNA二重鎖切断の修復経路です。正確な修復ではなく、つなぎ目に小さな欠失や挿入を残す「誤りがち」な仕組みで、DNAポリメラーゼθ(POLQ)が中心的に働きます。がんではこの経路への依存が新しい治療標的になり、CRISPRゲノム編集では逆にこの性質を「予測して利用する」応用が進んでいます。
- ➤正体 → HR・C-NHEJに次ぐ「第三の経路」。短い相同配列を利用して末端を結合する
- ➤主役の酵素 → DNAポリメラーゼθ(POLQ)。末端をつなぎ、ギャップを埋め、特徴的な変異を残す
- ➤最も活躍する場面 → 主要2経路が停止する有糸分裂(M期)で「最後の砦」として働く
- ➤がん治療 → BRCA変異がんはMMEJに依存。POLQを止めると選択的に死ぬ「合成致死」
- ➤ゲノム編集 → 修復結果を予測するinDelphi、短い配列で挿入するPITChに応用
1. MMEJとは:DNA修復の「第三の経路」
私たちの細胞は毎日、無数のDNAの傷にさらされています。その中でも、DNAの二本の鎖が同じ場所で切れてしまう「DNA二重鎖切断(DSB)」は、もっとも毒性の高い損傷です。放置すれば細胞死やがん化につながるため、細胞はこれを直すための修復経路をいくつも用意しています。詳しくはDNA修復のしくみもあわせてご覧ください。
💡 用語解説:DNA二重鎖切断(DSB)とは
DNAは、はしごをねじったような二本鎖の構造をしています。その二本が同じ位置でプツンと切れてしまうのがDNA二重鎖切断(Double-Strand Break: DSB)です。片方の鎖だけが切れた傷は「もう片方を見本にして直す」ことができますが、二本とも切れると見本になる相手がその場に無いため、修復はとても難しくなります。うまく直せないと、細胞死や、染色体がバラバラになる「ゲノム不安定性」を招き、がんの温床になります。
DSBを直す代表的な経路は、長らく次の2つと考えられてきました。ひとつは相同組換え(HR)で、姉妹染色分体という「まったく同じ配列のコピー」を見本に使うため、極めて正確に直せます。もうひとつが古典的非相同末端結合(C-NHEJ)で、見本を使わず切れた端どうしを素早くつなぐ、細胞周期を問わず働く経路です。ところが、これら2つの主要因子を欠いた細胞でも、DNAの末端はなお低い頻度でつながることが酵母や哺乳類の研究から見つかりました。そのつなぎ目には、数塩基の共通配列を伴う欠失が残っていたのです[1]。
こうして浮かび上がった「第三の経路」がMMEJ(マイクロホモロジー媒介性末端結合)です。代替的非相同末端結合(Alt-NHEJ)とも呼ばれ、切れたDNAの両端にたまたま存在する1〜25塩基ほどの極めて短い共通配列(マイクロホモロジー)を目印に、末端どうしを合わせてつなぎます[1]。この「短い相同配列を利用する」点が名前の由来です。
💡 用語解説:マイクロホモロジー(微小相同配列)
マイクロホモロジーとは、DNA配列の中に偶然できている「ごく短い、そっくりな並び」のことです。たとえば切れた左端に「GATTC」があり、右端の少し内側にも「GATTC」があれば、この2か所を貼り合わせて末端をつなぐことができます。HRが使う見本(姉妹染色分体まるごと)と比べると、MMEJが頼りにする手がかりはわずか数塩基と圧倒的に短いのが特徴です。短い手がかりで無理につなぐため、間の配列が失われて「欠失」が生じます。
MMEJの最大の特徴は、その「誤りがち(エラープローン)」な性質です。マイクロホモロジーを探す過程で末端が削られるため、修復後のつなぎ目には必ず欠失が残ります。さらに、後述する特殊なポリメラーゼの働きによって小さな挿入も加わりやすく、結果として染色体の転座・逆位・コピー数変異(CNV)といった複雑なゲノム再構成の原因になります[1]。一見すると危険なだけの仕組みに思えますが、実際には、HRが使えない状況で染色体の崩壊を食い止める高度に制御された生存戦略であることが分かってきました。
2. 分子メカニズム:POLQを中心とした精密装置
🔍 関連ページ:POLQ遺伝子/DNA損傷応答(DDR)/変異シグネチャー
MMEJは、C-NHEJの中心因子(Ku70/80やDNA-PKcsなど)にまったく頼らない独立した経路です。損傷の認識から末端の処理、貼り合わせ、DNA合成、そして最後のつなぎ止めまで、専門化した酵素がリレー形式で協調して働きます。その流れを図で示します。
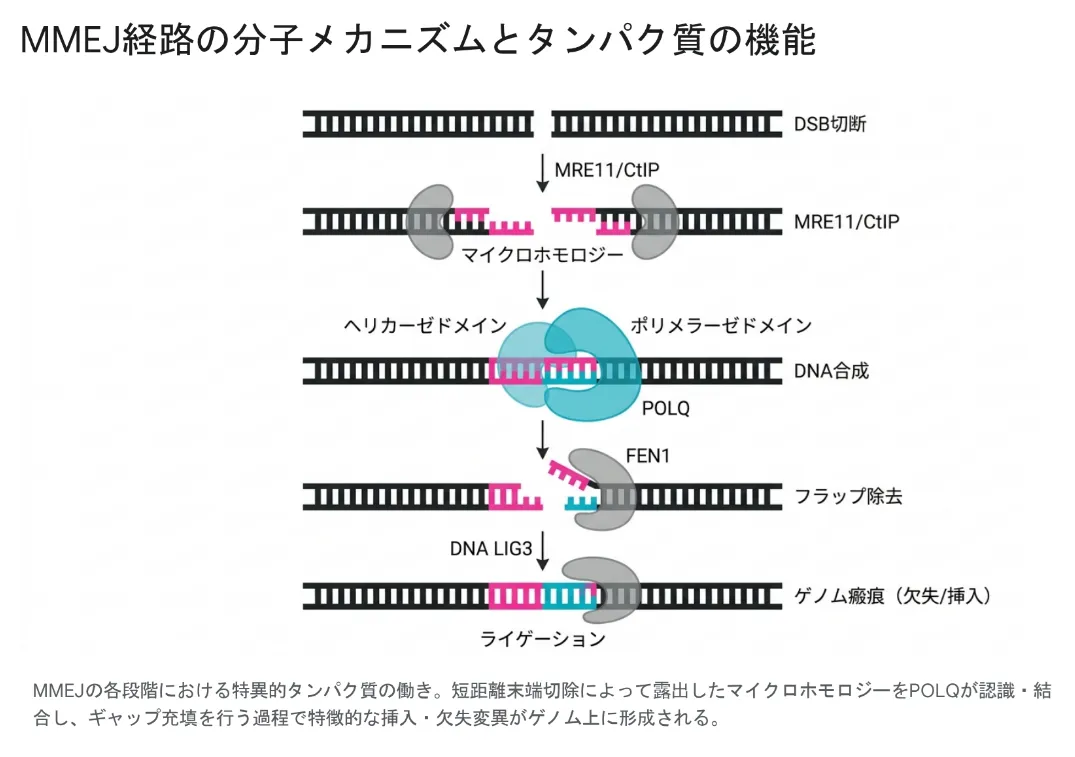
MMEJの各段階。MRE11/CtIPによる末端切除でマイクロホモロジーが露出し、POLQが認識・結合してギャップを埋め、FEN1が余分な配列を切り、DNA LIG3がつなぎ止める。つなぎ目には特徴的な欠失・挿入が残る。
ステップ①:短距離の末端切除で手がかりを露出させる
MMEJの最初の一歩は、DSBの端を少しだけ削る「末端切除」です。C-NHEJが端をそのまま結ぼうとするのに対し、MMEJはMRE11ヌクレアーゼやCtIPという因子による切除を必須とします。この切除で、ふだんは二本鎖の内側に隠れているマイクロホモロジーが一本鎖として露出します。哺乳類細胞では、Ku70が無い状況でも、この短距離切除(20塩基未満)にMRE11の酵素活性が欠かせないことが確認されています[2]。
重要なのは、切除の「長さ」です。HRの場合はExo1やBLMヘリカーゼによって数百〜数千塩基におよぶ「長距離切除」が行われますが、長すぎる切除はむしろMMEJを妨げます。MMEJは、ごく短い切除の段階で素早く次のステップへ移る、いわば「限られた切除条件に最適化された経路」なのです[2]。
ステップ②:DNAポリメラーゼθ(POLQ)が貼り合わせて埋める
💡 用語解説:DNAポリメラーゼθ(POLQ/Polθ)
DNAポリメラーゼθは、MMEJの主役となる特殊な酵素です。片方の端に「DNAをほどく・貼り合わせるヘリカーゼ部分」、もう片方の端に「DNAを合成するポリメラーゼ部分」を持つユニークな構造をしています。動物や植物では、MMEJは事実上このPOLQに依存しており、そのため「POLθ依存的末端結合(TMEJ)」とほぼ同じ意味で使われるほどです。POLQは、わずか1塩基以上のごく短いマイクロホモロジーを探して端を合わせ、隙間(ギャップ)を埋める合成を行います。
POLQのポリメラーゼ活性は、通常のDNAポリメラーゼとは大きく異なります。ふつうのポリメラーゼは正確な塩基対(AとT、GとCの組み合わせ)がそろわないと合成を始められませんが、POLQはミスマッチを含む不完全な組み合わせからでも合成を開始できます[4]。このため、MMEJは単に端を削ってつなぐ(欠失を残す)だけでなく、つなぎ目に小さなDNA配列を差し込む「鋳型依存性挿入」を高い頻度で起こします。
鋳型依存性挿入:「スナップバック」と「ループアウト」
この特徴的な挿入は、POLQが合成の途中で見本にする鎖を頻繁に乗り換える「すべりやすさ」に由来します。主に2つのモデルで説明されます。ひとつは「スナップバック機構」で、切れた端の一本鎖が自分自身で折り返してヘアピン構造をつくり、自分のすぐ近くの配列を見本にして逆向きの相補配列を合成する現象です[4]。もうひとつは「ループアウト(トランス)機構」で、向かい側の別の鎖や、まったく別のゲノム領域に一時的に貼りついて配列をコピーしてから、元の鎖に戻ってつなぎを完成させます[5]。
こうした「合成→はがれる→貼り直す」の繰り返しによって、がんゲノムにPOLQ特有の変異シグネチャー(ゲノムに残る傷跡のパターン)が刻まれます。このシグネチャーは、報告によって幅はあるものの、非常に多くのがん種で検出され、Polθによる修復の痕跡と考えられています[10]。反応速度が遅いために一本鎖DNAが長く露出しやすく、それが過剰な変異生成につながる可能性も指摘されています[3]。
ステップ③:FEN1が余分を切り、LIG3がつなぎ止める
POLQによる貼り合わせと合成のあとには、完全に相補的にならずに余ってしまったDNA配列(フラップ)が生じます。この余分な部分はFEN1(フラップ・エンドヌクレアーゼ1)という酵素が正確に切り取ります。最後にDNAリガーゼIII(LIG3)が残った切れ目(ニック)を封じて、一連の修復が完了します[3]。このように、MMEJは複数の専門化した酵素が順にバトンを渡す精密なリレーとして進みます。
3. 4つの修復経路を比較する
DSBを直す経路は、必要とする「見本の長さ」「正確さ」「主に働く細胞周期」がそれぞれ異なります。MMEJの位置づけを理解するために、代表的な4経路を並べて比較してみましょう。一本鎖アニーリング(SSA)は、より長い相同配列を使って大きな欠失を残す経路で、MMEJと似ていますが必要な相同配列の長さが異なります。
この表から、MMEJは「HRほど正確ではないが、C-NHEJより長い相同配列を利用し、SSAほどは長くない」という中間的な立ち位置にあることが分かります。正確さでは劣るものの、他の経路が使えない場面で確実に末端を結べる点に、この経路の存在意義があります。
4. 細胞周期による制御とM期の「最後の砦」
どの修復経路が働くかは、細胞周期(細胞が分裂に向けて進む一連の段階)と密接に結びついています。MMEJは常に一定の力で働いているわけではなく、特定の段階でこそ重要になるよう、厳密に制御されています。細胞がほとんど分裂していないG1期ではMMEJの活性は非常に低く、この時期のDSB修復はC-NHEJが担います。ところが、DNAを複製するS期に入るとMMEJの活性が高まります。複製の途中でフォーク(複製の進行点)が壊れて生じるDSBを処理する際、C-NHEJとHRの両方が抑えられている状況では、MMEJが数少ない選択肢になり得るためです[3]。
細胞周期の進行に伴う主要DNA修復経路の相対的な活性
概念図:値が大きいほど、その時期に活発に働くことを示す
G1期
S期
G2期
M期(有糸分裂)
G1期はC-NHEJが主導し、S/G2期ではHRが活性化します。特筆すべきはM期で、主要経路が抑えられる中、MMEJがバックアップ修復として単独で機能します。
そして近年の画期的な発見が、MMEJが最も不可欠になるのは有糸分裂(M期)だということです。細胞が2つの娘細胞に分かれるM期には、染色体がぎゅっと凝集します。この時期、HRとC-NHEJは、テロメア融合のような無秩序な結合を防ぐためにあえて完全にスイッチオフされています。もしS期やG2期のDSBが直りきらないままM期に持ち越されると、染色体の断裂という破滅的な危機に直面します。
この脆弱なM期に、細胞を救う緊急バックアップとして働くのがMMEJです。しかもこの動きは無作為ではなく、「RHINO」というタンパク質が鍵を握ります。RHINOはM期に蓄積し、細胞周期の調節因子であるPLK1(Polo様キナーゼ1)によってリン酸化を受けます。修飾されたRHINOがPOLQと結びついて、未修復のDSBへPOLQを直接呼び込むのです[9]。こうしてM期に強力なMMEJが引き起こされ、持ち越された切断が娘細胞へ分配される前に、いわば強引にでも修復されます。

5. 修復経路のクロストークとPARP1の役割の再定義
DNA修復経路は、それぞれ独立に動いているわけではありません。互いに競合し、複雑なやりとり(クロストーク)を通じて「どの経路で直すか」という経路選択が決まります。とりわけMMEJとHRは、密接な拮抗関係にあります。
末端切除が始まると、まずRPA(Replication Protein A)という因子が一本鎖DNAに素早く結合し、マイクロホモロジーどうしの不適切な貼り合わせを立体的に邪魔します。その後、BRCA2の助けを借りてRAD51がRPAと置き換わり、細胞をHRの方向へ強く導きます[7]。つまり、HRの因子が正常に働いている限り、変異を伴うMMEJは「安全装置」で抑え込まれているのです。ところが、高感度な変異解析を用いた研究では、BRCA1・BRCA2・RPAを枯渇させた細胞では、MMEJに特徴的な2〜6塩基のマイクロホモロジーを介した欠失変異が劇的に増えることが示されました[7]。HRに欠陥が生じると、MMEJが抑制を逃れ、変異を伴いながらDSB修復を肩代わりするのです。
PARP1をめぐるパラダイムシフト
🔍 関連ページ:PARP阻害剤とは/PARPトラッピング/PARP1遺伝子
MMEJの開始に関しては、近年大きな認識の転換が起きています。長年、PARP1はMMEJを初期段階で後押しする必須因子で、POLQを呼び込むのに欠かせないと考えられてきました。ところが、修復経路が無傷の細胞を使った最新の系統的研究により、この定説が覆されつつあります。細胞周期を厳密に制御した実験で、さまざまなPARP阻害剤を加えると、POLQ依存的なMMEJが下がるどころかむしろ「上昇」したのです[8]。
この現象は次のように説明されます。PARP1(PARP2ではありません)の阻害は、特定の切断部位でHRなどの競合経路による修復を妨げます。その結果、修復の経路選択が変わり、POLQ依存的なMMEJへの依存度が高まるのです[8]。さらに決定的なことに、MMEJが支配的となるM期では、PARP1もPARP2も修復に「不要(dispensable)」であることが示されました。POLQを止めるとM期のDSBが未修復のまま持ち越されて小核が増えるのに対し、PARP阻害剤(オラパリブなど)はこの小核形成にほとんど影響を与えませんでした[8]。
📝 この発見の臨床的な意味:「PARP阻害剤を使うとMMEJも一緒に抑えられてしまうのでは」という懸念がありましたが、実際にはPARP阻害下でもMMEJは働き続けます。これは、PARP阻害剤とPOLQ阻害剤を併用してがん細胞を追い込む治療戦略の、強い理論的裏づけになりました[8]。
6. がん治療への応用:合成致死性とPOLQ阻害剤
🔍 関連ページ:合成致死性/相同組換え修復欠損(HRD)/POLQ遺伝子
MMEJは染色体の転座や欠失を招くため、多くのがんの土台となるゲノム不安定性の主要な推進役です。実際、慢性骨髄性白血病や膀胱がんの一部では、C-NHEJ因子が減る一方でMMEJ因子が増え、DSB修復をMMEJに大きく依存していることが観察されています[3]。この極端な依存が、特定のがん細胞の「アキレス腱」になり得ます。
BRCA変異がんにおける「合成致死性」
💡 用語解説:合成致死性(Synthetic Lethality)
合成致死性とは、2つの遺伝子のうち片方だけが壊れても細胞は生きられるのに、両方が同時に壊れると細胞が死ぬ、という関係のことです。がん治療では、がん細胞ですでに壊れている遺伝子(例:BRCA)と「合成致死」の関係にある別の遺伝子を薬で狙い撃ちすると、正常細胞は生き残り、がん細胞だけが死ぬ——という選択的な攻撃が可能になります。BRCA変異がんに対するPARP阻害剤が、この考え方の代表例です。
乳がんや卵巣がんの多くは、BRCA1またはBRCA2の変異を持ち、HRが働かない状態、すなわち相同組換え修復欠損(HRD)に陥っています。正常細胞はHRが使えるため、MMEJを止められてもDSBを正確に直して生き延びます。しかしHRを失ったBRCA変異がん細胞は、複製中の切断やM期の未修復DNAを処理するのにMMEJ(特にPOLQ)へ完全に依存しています[6]。この状況でPOLQを止めると、DSBがまったく直せなくなり、染色体の粉砕と細胞死が引き起こされます。これがPOLQ阻害による合成致死のしくみです。
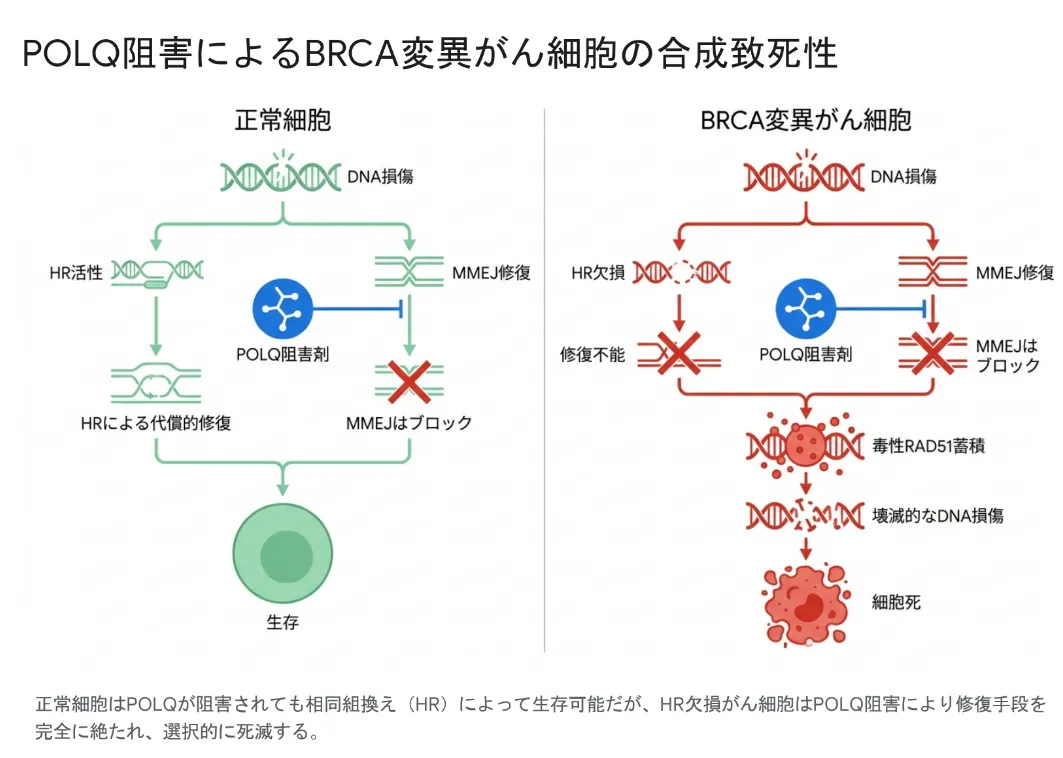
正常細胞はPOLQを阻害されても相同組換え(HR)で生存できるが、HR欠損がん細胞はPOLQ阻害で修復手段を完全に絶たれ、選択的に死滅する。
さらにPOLQ阻害は、単に修復を止めるだけではありません。POLQは本来、切除された一本鎖DNA上にRAD51が過剰にたまるのを防いでいます。POLQを止めるとこの制御が外れ、一本鎖DNA上にRAD51が毒性を伴って異常蓄積し、がん細胞の死をさらに加速させます[6]。POLQは正常組織ではほとんど発現しない一方、多くのがんで強く過剰発現しているため、がんに特異的な理想的標的として注目されています[6]。
POLQ阻害剤の開発と併用療法
この強固な根拠に基づき、POLQ阻害剤の開発が世界中で急速に進んでいます。もともと抗生物質として知られていたノボビオシンが、POLQのヘリカーゼ部分を狙う阻害剤として働き、生体内でHR欠損腫瘍を選択的に死滅させることが見いだされました[6]。近年はより強力で特異的な低分子化合物(ART558・ART4215・ART6043・RP-6685・SYN818など)が臨床段階へ進み、HR修復欠損の固形腫瘍を対象としたAZD4956の第1/2a相試験(PARTHENON試験)なども進行しています[17]。
単剤だけでなく、併用療法の研究も活発です。PARP阻害剤に耐性を得た腫瘍は、生存をいっそうMMEJに頼るようになる傾向があるため、POLQ阻害剤とPARP阻害剤の併用が、耐性を突破する相乗効果として期待されています[6]。前のセクションで述べた「PARP阻害下でもMMEJが働き続ける」という知見[8]は、この併用戦略の合理性をいっそう裏づけています。ほかにも、複製ストレスに関わるATR-CHK1-WEE1経路の阻害剤や、FEN1・DNA-PKを標的とする薬剤との組み合わせ、放射線療法との併用による放射線増感なども研究されています[6]。
7. ゲノム編集への革新的応用
🔍 関連ページ:CRISPR-Cas9/ゲノム編集とは
MMEJの「短い配列に依存して、決まった欠失や挿入を生む」という性質は、病気の原因変異を説明するだけでなく、人為的にゲノムを書き換える「精密ゲノム工学」に革新的な道具をもたらしました。CRISPR-Cas9などでDSBを入れて細胞自身の修復に委ねると、以前は多様な挿入欠失が無秩序に生じる「ノイズの多い」過程と考えられていました。
修復結果を予測する:inDelphiとMENdel
ところが、MMEJのしくみを計算科学的に解析した結果、この認識は覆されました。MMEJは切断部位の周辺配列とマイクロホモロジーの重なり長で修復パターンが決まるため、生じる挿入欠失を高い精度で予測できるのです。この原理を機械学習に落とし込んだのが「inDelphi」などのモデルです。inDelphiは、細胞株において配列情報だけから編集産物の80〜95%の挿入欠失長と、65〜80%の遺伝子型を正確に予測できることが示されました。さらに、特定のガイドRNAの5〜11%では、生じる編集産物の半分以上が単一の均一な遺伝子型になる「precise-50」という状態が起こることも分かりました[11]。この性質を利用すると、修復の見本となるドナーDNAを一切入れずに、細胞自身のMMEJだけで、195ものヒト疾患関連の配列を高い再現性で健康な配列へ「修正」できる可能性が示されています[11]。
大規模な検証では、MMEJによる均一な遺伝子型を見つける感度において、MENTHUというアルゴリズムがinDelphiやLindelを上回ることが報告されました。これらを統合した「MENdel」は、脊椎動物のコード領域で狙える均一なフレームシフト変異の頻度を高い精度で見積もることを可能にし、遺伝子機能の研究や、機能喪失を利用した遺伝子治療応用への道を開いています[12]。
PITChシステム:極小の相同配列で外来遺伝子を挿入する
MMEJのもうひとつの応用が、外来遺伝子を組み込む「ノックイン」です。従来のノックインはHRに依存し、数百〜数千塩基もの長い相同配列(ホモロジーアーム)を持つドナーが必要で、作製が煩雑でした。しかもHRはS/G2期でしか働かないため、分裂を止めた細胞では効率が落ちます。この問題を解決したのが、日本の研究チームが開発した「PITChシステム」です。MMEJが5〜25塩基程度の短い相同配列で機能することを利用し、ドナーに付ける相同配列をわずか10〜40塩基で済ませます。この長さならPCRプライマーに配列を含めるだけで付加でき、数日でドナーを作製できます[13]。余分な配列がMMEJの過程で自動的に削り取られるため、継ぎ目のない精密なノックインを実現できる点が大きな強みです[13]。
生物種を超えたMMEJの普遍性
MMEJは哺乳類や酵母だけでなく、多様な生物に広く保存されています。ゼブラフィッシュの胚発生では、POLθやDNAリガーゼ3を働かなくしても発生自体は妨げられないものの、放射線への感受性が高まり、Cas9による修復時の変異パターンが大きく変わることが示されました[15]。一方、蚊(ネッタイシマカ)ではCRISPRで生じたDSB修復の大半をMMEJが担っており、遺伝子ドライブ戦略の設計に重要な知見となっています[14]。こうした違いは、生物種や発生段階に応じて各経路が役割分担しながらゲノムを守っていることを物語っています。酵母の研究では、長い非相同末端の除去にRad1-Rad10ヌクレアーゼが関わることも古くから知られています[16]。
8. 遺伝子診断・遺伝カウンセリングとのつながり
🔍 関連ページ:BRCA遺伝子検査/遺伝性乳がん卵巣がん症候群(HBOC)/臨床遺伝専門医とは
MMEJは分子生物学の基礎概念ですが、遺伝診療との接点は明確です。ここまで見てきたとおり、HRDを示すBRCA変異がん(HBOCなど)では、がん細胞がMMEJ(POLQ)に生存を頼っています。つまり「どの修復経路が壊れているか」という遺伝情報が、そのまま「どの薬が効きやすいか」という治療選択の手がかりになります。この意味でMMEJは、遺伝性腫瘍の遺伝子診断から治療戦略へつながる橋渡しの概念だといえます。
こうした変異特異的な治療を検討する前提として、まず原因となる遺伝子変異を正確に同定することが欠かせません。BRCA遺伝子検査などで変異の有無やその意義を評価し、その結果をどう理解し、家族にどう共有するかを、臨床遺伝専門医による遺伝カウンセリングで丁寧に扱います。検査結果は本人だけでなく血縁者の健康管理にも関わるため、専門的な支援のもとで意思決定を進めることが大切です。
一方で、inDelphiやPITChに代表されるゲノム編集への応用は、現時点では研究段階・遺伝子治療研究の基盤という位置づけです。MMEJを利用した治療がすぐに日常診療で使えるわけではありません。本記事は、いま世界の研究で何が起きているのかを正確にお伝えするための学術的な解説であり、特定の検査や治療をおすすめするものではありません。
9. よくある誤解
誤解①「MMEJはただの予備でしかない」
かつてはそう考えられていましたが、いまでは主要2経路が止まるM期に細胞を救う不可欠な経路と分かっています。単なるバックアップではなく、特定の場面では主役として働きます。
誤解②「誤りがちだから、悪者にすぎない」
確かにMMEJは欠失や挿入を残しますが、それは染色体崩壊という最悪の事態を避けるための代償です。細胞にとっては「傷跡は残っても生き延びる」ための合理的な選択でもあります。
誤解③「PARP阻害剤を使えばMMEJも止まる」
最新研究では逆で、PARP阻害下でもMMEJはむしろ活発になることが示されました。だからこそPOLQ阻害剤との併用に意義があると考えられています。
誤解④「POLQ阻害剤はもう使える治療だ」
POLQ阻害剤の多くは臨床試験・研究段階です。有望な戦略ですが、標準治療として確立されたものではなく、当院で提供する治療でもありません。
よくある質問(FAQ)
🏥 遺伝性腫瘍・遺伝子診断のご相談
BRCAをはじめとする遺伝性腫瘍の遺伝子検査や
遺伝カウンセリングは、臨床遺伝専門医が在籍する
ミネルバクリニックにお気軽にご相談ください。
参考文献
- [1] Microhomology-mediated end joining: a back-up survival mechanism or dedicated pathway? Trends Biochem Sci (PMC). [PMC4638128]
- [2] Microhomology-mediated End Joining and Homologous Recombination share the initial end resection step to repair DNA double-strand breaks in mammalian cells. PNAS. [PNAS]
- [3] Microhomology-mediated end joining: Good, bad and ugly. Mutat Res (PMC). [PMC6477918]
- [4] Mechanism of Suppression of Chromosomal Instability by DNA Polymerase POLQ. PLOS Genetics. [PLOS Genetics]
- [5] Risky Business: Microhomology-Mediated End Joining. Mutat Res (PMC). [PMC4887395]
- [6] Exploiting the Microhomology-Mediated End-Joining Pathway in Cancer Therapy. Cancer Res (AACR). [AACR Cancer Res]
- [7] A role for human homologous recombination factors in suppressing microhomology-mediated end joining. Nucleic Acids Res (PMC). [PMC4937322]
- [8] PARP1 and PARP2 are dispensable for DNA repair by microhomology-mediated end-joining at double-ended DSBs. Nucleic Acids Research. 2026;54(1):gkaf1437. [NAR]
- [9] RHINO directs MMEJ to repair DNA breaks in mitosis. Science (PMC). [PMC10561558]
- [10] Defining the mutation signatures of DNA polymerase θ in cancer genomes. NAR Cancer. [NAR Cancer]
- [11] Predictable and precise template-free CRISPR editing of pathogenic variants. Nature (PMC). [PMC6517069]
- [12] Deploying MMEJ using MENdel in precision gene editing applications for gene therapy and functional genomics. Nucleic Acids Res (PMC). [PMC7797032]
- [13] Establishment of expanded and streamlined pipeline of PITCh knock-in — a web-based design tool for MMEJ-mediated gene knock-in. Bioengineered (PMC). [PMC5470537]
- [14] Microhomology-mediated end joining is the predominant form of DNA repair in the mosquito Aedes aegypti. PMC. [PMC12809539]
- [15] Dynamic interplay of cNHEJ and MMEJ pathways of DNA double-strand break repair during embryonic development in zebrafish. PMC. [PMC11811205]
- [16] Microhomology-Mediated End-Joining Chronicles: Tracing the Evolutionary Footprints of Genome Protection. PMC. [PMC12426944]
- [17] Study of AZD4956 (POLθ inhibitor) as Monotherapy and in Combination in HRR-Deficient Solid Tumours (PARTHENON). ClinicalTrials.gov. NCT07446855. [ClinicalTrials.gov]






