目次
- 1 1. クロモスリプシス(染色体粉砕)とは?がんゲノム進化の常識を覆した大発見
- 2 2. クロモスリプシスを定義する4つの分子生物学的特徴
- 3 3. 【発生メカニズム1】分配エラーによる「小核」の形成と核膜の崩壊
- 4 4. 【発生メカニズム2】ゲノムを粉砕するヌクレアーゼ「N4BP2」の特定
- 5 5. 【発生メカニズム3】無秩序な再結合とecDNA(細胞外環状DNA)の誕生
- 6 6. がんにおける臨床的意義とTP53異常との相乗的悪循環
- 7 7. 腫瘍微小環境と免疫応答の交絡:cGAS-STING経路のパラドックス
- 8 8. 生殖細胞系列におけるクロモスリプシスと先天性疾患
- 9 9. 奇跡の自然治癒:WHIM症候群が教える生命の可塑性
- 10 10. 最先端の検出技術:ShatterSeekアルゴリズムからAI診断まで
- 11 11. 次世代の治療標的としての可能性と未来への展望
- 12 よくある質問(FAQ)
- 13 参考文献
- 14 関連記事

📍 クイックナビゲーション
長年にわたり、がん細胞のゲノムは「少しずつ時間をかけて進化する」と考えられてきました。しかし、2011年に発見されたクロモスリプシス(染色体粉砕)は、この常識を根底から覆しました。単一の壊滅的なイベントによって染色体が粉々に砕け散り、無秩序に再構成されるこの現象は、全ヒトがんの約4分の1で観察される普遍的なメカニズムであり、がんの急速な悪性化や治療抵抗性に直結します。本記事では、染色体が粉砕される驚くべき分子メカニズムから、先天性疾患への影響、そして奇跡的な自然治癒の事例まで、最新の知見を臨床遺伝専門医が徹底的に解説します。
Q. クロモスリプシスとは一言で言うと何ですか?
A. 細胞分裂の際に生じたエラーによって特定の染色体が孤立し、細胞自身の酵素によって粉々に砕かれ、その後無秩序に再結合される現象です。この結果、遺伝子の配列が極めて複雑に入り乱れ、がん細胞ではがん遺伝子の異常な増幅やがん抑制遺伝子の喪失を引き起こして急速な悪性化を促します。
- ➤発生の引き金 → 分配エラーによる「小核」の形成と、その脆弱な核膜の崩壊。
- ➤直接的な破壊要因 → 細胞質に存在するヌクレアーゼ「N4BP2」等による無数のDNA切断。
- ➤がんにおける影響 → TP53異常との悪循環により、予後を著しく悪化させる。
- ➤先天性疾患と奇跡 → 生殖細胞系列でも発生。稀にWHIM症候群のように自然治癒をもたらすことも。
- ➤検出と未来 → 全ゲノムシークエンス(WGS)とShatterSeekによる検出、N4BP2阻害などの治療展望。
1. クロモスリプシス(染色体粉砕)とは?がんゲノム進化の常識を覆した大発見
長年にわたり、腫瘍学および遺伝学の分野において、がんのゲノムは「長い時間をかけて徐々に進化するもの」と考えられてきました。この古典的な「多段階発がんモデル」は、ダーウィンの漸進的進化論に基礎を置き、点突然変異や遺伝子の微小な欠失、あるいは小規模な重複といった遺伝的変異が、細胞分裂のたびに少しずつ蓄積することで、最終的に正常細胞が悪性化するというプロセスを提唱していました [1]。
しかし、この生物学における長年のドグマ(定説)は、2011年に報告された画期的な発見によって根底から覆されることとなりました。Stephens博士らの研究チームは、慢性リンパ性白血病(CLL)患者の全ゲノムを詳細に解析する過程で、従来の理論では到底説明のつかない、異常なゲノム変異のパターンを特定したのです [1]。
💡 用語解説:断続平衡説(Punctuated equilibrium)
進化は長期間の「安定期」と、短期間の「急激な変化」の繰り返しで起こるという進化生物学の理論です。クロモスリプシスの発見は、がん細胞のゲノム進化においても、徐々に変異が溜まるだけでなく、ある日突然、破局的な大イベントによって一気にゲノムが書き換わる「断続平衡説」が成立することを証明しました。
彼らが直面した事例では、一人の患者の「4番染色体長腕(4q)」という極めて限定された領域内に、なんと42箇所にも及ぶ大規模な構造変異(ゲノム再構成)が集中して発生していました。もしこれらの変異が古典的な理論に従って「徐々に」蓄積したとすれば、変異はゲノム全体にランダムに散在するはずです。単一の染色体の腕の部分にのみ、これほど高密度に集中することは、確率論的にあり得ないことでした [1]。
この観測結果から導き出された結論は、ゲノムの急速な不安定化を引き起こす「爆発的なメカニズム」が存在するというものでした。すなわち、細胞分裂の歴史における「単一の壊滅的なイベント(Single catastrophic event)」によって染色体が物理的に粉々に砕け散り、その後、細胞が生き延びるためのDNA修復機構によってランダムかつ無秩序に再結合されたというモデルです [1]。
この破壊的な現象は、ギリシャ語で「染色体」を意味する「khrôma」と、「粉々に砕けること」を意味する「thripsis」を組み合わせて、「クロモスリプシス(Chromothripsis:染色体粉砕)」と命名されました。現在では、この現象は稀な例外ではなく、全ヒトがんの約4分の1(25%)で観察され、特定の腫瘍においてはさらに高い頻度で発生する広範かつ普遍的な突然変異現象であることが明らかになっています [1]。
2. クロモスリプシスを定義する4つの分子生物学的特徴
🔍 関連記事:コピー数変異(CNV)/全ゲノムシークエンス(WGS)
クロモスリプシスが発見されて以降、この現象を他の複雑なゲノム再構成から区別し、正確に同定するための厳密な分子基準の確立が求められてきました。研究者のHollandとClevelandは、複雑な染色体再構成を生み出す破局的イベントの総称として「クロモアナジェネシス(Chromoanagenesis)」という包括的な概念を提唱し、クロモスリプシスをその特定のサブセットとして明確に位置付けました。
国際ヒト細胞ゲノム命名規約(ISCN)のガイドラインにおいて、クロモスリプシスは「染色体または染色体セグメントの長さに沿って、遺伝子コピー数の変化(正常、獲得、または喪失)が交互に現れる複雑なパターン」として定義されています。この現象を特徴付ける具体的な4つの基準は以下の通りです。
① クラスター化されたDNA二重鎖切断(Clustered Breaks)
影響を受けたゲノム領域には、互いに極めて近接した多数のDNA切断が密集して存在する「クラスター領域」が形成されます。重要なことは、これらの破壊されたクラスター領域の周囲には、再構成の影響を全く受けていない無傷の巨大なDNA領域が広がっている点です。これは、破壊がゲノム全体ではなく、特定の局所に限定された壊滅的イベントであることを示しています。
② コピー数の急速な振動(Oscillation of Copy Number States)
粉砕された染色体が再構築される際、一部のDNA断片は完全に失われ(欠失)、残された断片だけで不完全な修復が行われます。その結果、ゲノムのコピー数が「正常(多くの場合2コピー)」と「喪失(1コピーまたは0コピー)」の間で交互かつ頻繁に変動するプロファイルを示します。特筆すべきは、クロモスリプシスにおいては局所的な喪失が主であり、重複や三重重複といった高次の増幅を伴わないという点です。
③ 単一ハプロタイプの関与(Single Haplotype Involvement)
原則として、この破壊的イベントは一対の相同染色体(父親由来と母親由来)のうち、片方の親由来のコピー(単一ハプロタイプ)のみで発生します。両方が同時に粉砕されることは通常ありません。
④ 不完全かつランダムな再構成と配向の変化
修復プロセスを通じて断片が繋ぎ合わされる際、その順序はランダムになり、断片の向きが逆転する(逆位)などの構造的変動が多発します。また、再結合されなかった断片は消失するか、あるいは主たる染色体から独立して「ダブルマイナス」と呼ばれる細胞外環状DNA(ecDNA)を形成して存続することがあります。
表に示されるように、クロモアナシンセシスが「異常なDNA複製」に起因するのに対し、クロモスリプシスは純粋に「物理的な染色体の分断と再結合」に起因するという点が最も本質的な差異です。この鑑別は、次世代シーケンシングデータを解釈し、発がんの真のメカニズムを特定する上で極めて重要です。
3. 【発生メカニズム1】分配エラーによる「小核」の形成と核膜の崩壊
長年にわたり、細胞内でいかにして染色体が文字通り「粉砕」されるのか、その引き金となる物理的・化学的要因は最大の謎でした。ゲノム全体が無作為にダメージを受ける放射線被曝などとは異なり、クロモスリプシスは特定の染色体またはその一部領域にのみ限定して発生します。この空間的な特異性を説明するメカニズムとして特定されたのが、細胞分裂時のエラーに起因する「小核(Micronucleus)」の形成と、その崩壊プロセスです [2]。
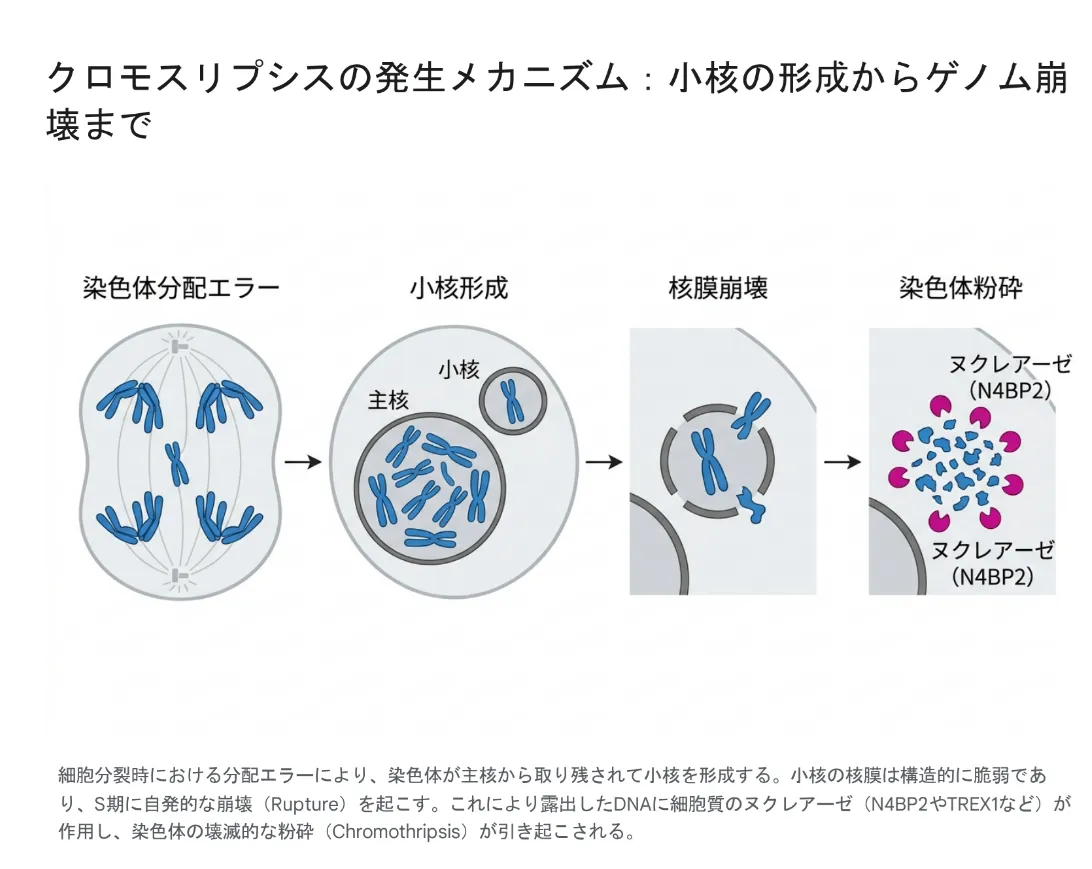
図:クロモスリプシスの発生メカニズム。小核の形成から、脆弱な核膜の崩壊、そして細胞質ヌクレアーゼによるゲノム崩壊へのカスケード。
染色体分配エラーによる隔離
細胞が有糸分裂(細胞分裂)を行う際、複製された染色体は紡錘体(Mitotic spindle)によって両極の娘細胞へと均等に分配されます。しかし、動原体の機能異常や分裂チェックポイントの欠陥が生じると、一部の染色体(特に遺伝子密度が低くサイズの大きい染色体など)が正常に極へと引き寄せられず、分裂の後期において「遅滞染色体」として取り残される事態が発生します [2]。
正常に分配された染色体群は新たな細胞の「主核(Primary nucleus)」を再構築しますが、取り残された遅滞染色体はこの主核には取り込まれません。その代わり、独自の小さな核膜に包まれた「小核」として細胞質内に独立して存在するようになります。この時点で、遅滞染色体は細胞の主要なゲノム保護環境から物理的に完全に隔離された状態となります [2]。
💡 用語解説:小核(Micronucleus)
正常な細胞のメインの核(主核)とは別に、細胞分裂時のエラーで細胞質に取り残された染色体が形成する「極めて小さな核」のことです。小核の存在自体が、細胞のDNAに傷がついていること(ゲノム不安定性)を示す強力なマーカーとして、がん細胞などで頻繁に観察されます。
核膜の極端な脆弱性と自発的崩壊
クロモスリプシスへの引き金となる小核の最大の特徴は、その「核膜(Nuclear Envelope)の極端な脆弱性」にあります。Salk研究所やHarvard大学の先駆的な研究により、小核の核膜がなぜ脆弱になるのかが分子レベルで解明されました。
主核が形成される正常な細胞では、核膜タンパク質が適切に集合します。しかし、小核を形成する遅滞染色体の場合、細胞分裂の糸である「微小管」と物理的に近接した位置で無理やり核膜形成が進行します。すると、この微小管自体が障害物となり、細胞質と核内の物質のやり取りをするドアの役割を果たす「核孔複合体」の組み立てを強力に阻害してしまうのです。
ドア(核孔複合体)が欠如した小核は、核膜の維持やDNA修復に不可欠なタンパク質を外部から取り込むことができません。この物資輸送の欠陥は、内部のDNA複製を著しく遅延・妨害させます。これらの構造的・機能的欠陥が蓄積した結果、細胞が次のDNA合成期(S期)に進行した直後、小核の核膜は耐えきれずに不可逆的な「自発的崩壊(Rupture)」を起こしてしまうのです [2]。
4. 【発生メカニズム2】ゲノムを粉砕するヌクレアーゼ「N4BP2」の特定
小核の核膜が崩壊すると、内部に隔離されていた染色体は細胞質の環境に直接曝露されます。細胞質には不要物を分解する様々な酵素が存在しますが、十数年にわたり「具体的にどの酵素が染色体を粉砕する直接的な引き金(Spark)となっているのか」は不明のままであり、クロモスリプシスを治療標的とすることを極めて困難にしていました。
破壊的ヌクレアーゼ「N4BP2」の発見
この長年の謎は、カリフォルニア大学サンディエゴ校(UCSD)のDon Cleveland研究室とKsenia Krupinaらの画期的な研究によって遂に解き明かされました。彼らは、既知および推定されるすべてのヒトヌクレアーゼ(DNA分解酵素)を対象とした大規模なスクリーニングを実施し、崩壊した小核内の染色体断片化を促進する酵素を探索しました [3]。
その結果、N4BP2と呼ばれるエンドヌクレアーゼが、崩壊した小核の内部に特異的に侵入し、露出したDNAを無数に切断する唯一の酵素であることが特定されたのです [3]。この発見を裏付ける決定的な証拠として、研究チームは脳腫瘍細胞からN4BP2を排除(ノックアウト)する実験を行いました。すると、N4BP2を取り除いた細胞では、染色体の粉砕が劇的に減少しました。
さらに反対に、N4BP2を人為的に「健康な細胞の主核内」に強制発現させると、本来無傷であるはずの正常な染色体であっても壊滅的に砕け散り、ゲノム崩壊が引き起こされることが確認されました [3]。
💡 用語解説:ヌクレアーゼ(Nuclease)
DNAやRNAを構成する核酸の鎖を切断・分解する酵素の総称です。鎖の途中を切るものをエンドヌクレアーゼ、端から削っていくものをエキソヌクレアーゼと呼びます。N4BP2は強力なエンドヌクレアーゼであり、通常は細胞質に存在しますが、小核の核膜が破れることで内部に侵入し、無防備なDNAを容赦なく切り刻みます。
この画期的な知見は、クロモスリプシスが単なる物理的ストレスや受動的な分解の蓄積ではなく、N4BP2という特定の酵素の能動的かつ過剰な切断活性によって引き起こされるカスケードであることを証明しています。実際に、10,000以上のヒトがんゲノムを解析した結果、N4BP2の発現レベルの上昇は、クロモスリプシスの発生およびそれに伴う細胞外環状DNA(ecDNA)の形成を強力に予測する因子であることが確認されています [3]。
TREX1とその他の関連因子
N4BP2に加えて、細胞質の主要なエキソヌクレアーゼであるTREX1も、クロモスリプシスに関与する酵素として注目されています。TREX1は通常、小胞体に局在し、細胞質に漏れ出した不要なDNAを分解して自己免疫応答を防ぐ役割を持ちますが、テロメア危機などの細胞ストレス下においてクロモスリプシスを媒介することが示唆されています。TREX1は崩壊した小核に動員されることで、内部のDNAを分解し粉砕をさらに促進する可能性があります。
特筆すべきは、真核生物の細胞質環境が、正常な染色体(主核に守られている)には影響を与えず、小核に取り残されたような「既存の欠陥を持つ染色体」を選択的に標的としてダメージを与えるという、合目的的な排除メカニズムを備えている点です。
5. 【発生メカニズム3】無秩序な再結合とecDNA(細胞外環状DNA)の誕生
🔍 関連記事:相同組換え修復
N4BP2やTREX1によって数千もの断片に粉砕されたDNAは、そのまま細胞質に放置されるわけではありません。細胞に備わった強力なDNA修復機構、主に「非相同末端結合(NHEJ:相同性を必要とせず壊れた末端同士を直接結合する機構)」などが働き、断片を強引に繋ぎ合わせようとします。しかし、本来一本の連続した糸であるはずの染色体が数千もの断片に分割されているため、元の正しい順序と向きで再構築されることは絶対に不可能です。
CIP2A-TOPBP1複合体による再統合の触媒
ライブセルイメージングと単一細胞ゲノム解析を組み合わせた戦略により、小核で粉砕された染色体が、次の細胞分裂の過程で、なんと「主核(Primary nucleus)」へと再統合(Reintegration)される驚くべき様子が捉えられています。この再統合プロセスを積極的に促進しているのが、CIP2A-TOPBP1タンパク質複合体です [4]。
CIP2Aは強力な腫瘍抑制因子を阻害する発がん性タンパク質であり、一方のTOPBP1はDNA複製や修復に関わる多機能な足場タンパク質です。小核内で染色体が粉砕された際、このCIP2A-TOPBP1複合体が断片化したDNAを物理的にクラスター化(束ねる)する働きを担います [4]。これにより、バラバラになった染色体断片が細胞質全体に散逸するのを防ぎ、次の細胞分裂における紡錘体の働きと同調して、主核へと効率的かつ忠実に持ち込むことが可能になるのです [4]。
発がんドライバーの形成:ecDNAと圧倒的な遺伝子増幅
主核へと再統合されたものの、全ての断片が染色体本体に繋ぎ合わされるわけではありません。再結合の過程から脱落した一部のDNA断片は、細胞内で端と端が結合して環状化し、「細胞外環状DNA(ecDNA:extrachromosomal DNA)」あるいはダブルマイナスと呼ばれる構造を形成します [3]。N4BP2の過剰発現によるクロモスリプシスは、このecDNAの生成を爆発的に促進することが確認されています [3]。
💡 用語解説:ecDNA(細胞外環状DNA)
通常の染色体(線状)から切り離され、細胞核内に独立して存在するリング状のDNA断片です。ecDNAは通常の染色体のように分裂時に均等に分配されないため、特定のがん細胞内で数が爆発的に増え(遺伝子増幅)、がんを極めて悪性化させる強力な原動力となります。
ecDNAはセントロメア(分配のための牽引ポイント)を持たないため、細胞分裂の際に娘細胞に対して均等に分配されず、ランダムに偏って受け継がれます(不均等分配)。これにより、特定の細胞系列においてecDNA上の遺伝子コピー数が劇的に増加し、数十から数百倍にも達することがあります。
もしこのecDNA上にMycなどの強力な「がん遺伝子(Oncogene)」が含まれていた場合、圧倒的な遺伝子増幅が引き起こされ、腫瘍の極めて攻撃的な増殖と、抗がん剤に対する強力な薬剤耐性の獲得へと直結するのです。クロモスリプシスは、単なる遺伝子の破壊にとどまらず、このようにしてがん細胞に新たな武器を与え、その生存を劇的に有利にするメカニズムとして機能しています。
6. がんにおける臨床的意義とTP53異常との相乗的悪循環
クロモスリプシスは、発見当初こそ一部の白血病や骨腫瘍における特殊な事例と考えられていましたが、WGSを用いた大規模な汎がん(Pan-cancer)解析プロジェクトの進展により、あらゆる臓器・組織の腫瘍において発生し得る普遍的なメカニズムであることが立証されました。
腫瘍タイプによる発生頻度の差異と影響
腫瘍のタイプによって、クロモスリプシスが構造変異を牽引する割合は大きく異なります。例えば、原発性および転移性の大腸がん(Colorectal cancer)においては、点突然変異や単純なコピー数変化と複雑に相互作用しながら、腫瘍の発生と転移を促進する主要なメカニズムとして機能していることが確認されています [5]。
また、ほぼすべての骨肉腫(Osteosarcoma)においてクロモスリプシスの痕跡が認められるほか、小児脳腫瘍であるソニック・ヘッジホッグ髄芽腫や神経芽腫でも極めて高い頻度で発生し、腫瘍の悪性度と深く結びついています。膵神経内分泌腫瘍(pNENs)の研究では、クロモスリプシスを持たない患者の生存期間中央値が2056日であったのに対し、有する患者では658日へと激減し、遠隔転移リスクが46%から92%に倍増することが報告されています。
TP53異常との相乗的悪循環
クロモスリプシスの発生と臨床的悪性度を語る上で絶対に欠かせないのが、TP53遺伝子の変異・欠失との極めて強固な関連性です。TP53はコードされるp53タンパク質が「ゲノムの守護神」として知られ、DNAに修復不可能な甚大なダメージが生じた際、細胞周期を停止させアポトーシス(細胞死)を誘導する中心的な役割を担います。
7. 腫瘍微小環境と免疫応答の交絡:cGAS-STING経路のパラドックス
ゲノムの不安定性は、がん細胞自身の増殖能力を高めるだけでなく、腫瘍周囲の微小環境(Tumor microenvironment)における宿主の免疫応答とも複雑な相互作用を引き起こします。クロモスリプシスの発生は、一見すると免疫系を活性化するように思われますが、実際には逆の現象を引き起こしています。
cGAS-STING経路の活性化
小核の核膜崩壊によって細胞質に大量のDNA断片が流出すると、本来はウイルスの侵入を感知する自然免疫センサーである「cGAS-STING経路」が、細胞質のDNAを検知し強力に活性化されます。理論上、この経路の活性化は炎症性シグナル伝達を誘発し、腫瘍浸潤リンパ球や細胞傷害性T細胞を動員して「免疫監視」を強化し、腫瘍細胞の排除を促進するはずです。加えて、ゲノムの大規模な再構成は多数のネオアンチゲン(新生抗原)を生み出し、これも免疫細胞を活性化する要因となるはずでした。
免疫逃避とパラドックスの実態
しかし、The Cancer Genome Atlas(TCGA)のデータセットを用いたマルチオミクス解析は、直感に反する驚くべきパラドックスを明らかにしました。クロモスリプシスを有する腫瘍グループは、持たないグループと比較して、総免疫細胞の浸潤スコアが有意に減少していることが判明したのです [7]。また、浸潤している免疫細胞の細胞傷害活性も著しく損なわれていました [7]。
臨床データにおいても、クロモスリプシスを持つ患者では抗腫瘍免疫効果が弱く、免疫チェックポイント阻害薬(免疫療法)に対する応答性が低いことが示されています [7]。この矛盾の背景には、がん細胞が有する高度な「免疫逃避(Immune evasion)メカニズム」が存在すると考えられています。
クロモスリプシスによる大規模な再構成は、細胞表面に目印を出すHLA因子の欠失や、免疫を抑制するサイトカインの異常増幅を同時に引き起こしている可能性が高いのです。さらにがん細胞は持続的なcGAS-STINGシグナルを逆手に取り、慢性的な炎症を通じて周囲を「免疫抑制的な微小環境(制御性T細胞の動員など)」へと書き換え、自らの生存を有利に導く適応を遂げていると推測されます。
8. 生殖細胞系列におけるクロモスリプシスと先天性疾患
🔍 関連記事:遺伝カウンセリングとは
クロモスリプシスは体細胞(がん細胞)において発生するだけでなく、頻度としては極めて稀であるものの、精子や卵子といった「生殖細胞系列(Germline)」においても発生し得ます。この場合、生じた変異は受精卵を通じて次世代の個体全体へと受け継がれ、複雑な先天性異常、発達遅滞、自閉症スペクトラム、知的障害などを引き起こす原因となります [8]。
均衡型再構成の優位性と「生存バイアス」
がんゲノムで見られるクロモスリプシスは、コピー数の「正常」と「喪失」が激しく振動する、極めて「不均衡型(Unbalanced)」の変異です。これは腫瘍抑制遺伝子の喪失が発がんに有利に働くためです。
一方、発達障害や先天性異常の患者さんの生殖細胞系列で観察されるクロモスリプシスは、がんのような規則的に振動するコピー数状態をほとんど示しません。過去の報告の多くは、コピー数の変動を全く伴わない完全に「均衡型(Balanced)」の再構成であるか、わずか数カ所の変化しか伴わないものでした [8]。
💡 用語解説:生存バイアス(選択圧)
胚発生(受精卵から胎児になる過程)において、多数の重要遺伝子が欠失した高度に不均衡なゲノムを持つ初期胚は、細胞分裂を正常に進めることができず早期に自然流産となります。結果として、妊娠を継続し出生に至ることができるのは、「遺伝子の量自体は正常に保たれたまま、染色体の順序だけが複雑に入れ替わった」均衡型のクロモスリプシスを持つ受精卵に偏るのです。これを強力な選択圧(生存バイアス)と呼びます。
均衡型のクロモスリプシスであっても、その断裂部位が重要な遺伝子のコード領域や制御領域を偶然分断してしまった結果として、発達遅滞などの疾患が発現します [8]。生殖細胞系列におけるリスクは、細胞周期やDNA修復遺伝子に先天的変異を持つ家系(Li-Fraumeni症候群など)において統計的に高いことが知られています。
9. 奇跡の自然治癒:WHIM症候群が教える生命の可塑性
クロモスリプシスは通常、致死的ながんや重篤な先天性疾患を引き起こす「破壊者」です。しかし、極めて特異な状況下において、この壊滅的な現象が奇跡的に「疾患を治癒」させる方向に働いた驚異的な臨床例が存在します。それが2015年に報告されたWHIM症候群の女性患者(WHIM-09)の事例です [9]。
WHIM症候群とは
WHIM症候群は、いぼ(Warts)、低ガンマグロブリン血症、反復性感染症、そして骨髄カテキシス(成熟した白血球が骨髄内に留まり末梢血に出られない状態)を特徴とする、極めて稀な複合免疫不全症です。この疾患は、ケモカイン受容体であるCXCR4遺伝子の機能獲得型変異によって引き起こされ、受容体のシグナルが異常に亢進することで白血球の異常な生着と滞留が生じます。
たった一つの細胞で起きた「自己遺伝子治療」
この30代の女性患者は、幼少期から重度の感染症に苦しんでいましたが、30代に入り突如として自然にすべての症状が消失しました。研究チームが彼女の血液とゲノムを詳細に解析した結果、彼女の骨髄内に存在する「たった一つの造血幹細胞(HSC)」において、2番染色体のクロモスリプシスが発生していたことが判明したのです [9]。
この染色体粉砕という壊滅的イベントによって、偶然にも変異を持ったCXCR4対立遺伝子(疾患アレル)を含む164の遺伝子群が一挙に欠失していました。なぜ、たった一つの幹細胞で起きたイベントが患者全体を治癒させたのでしょうか。
10. 最先端の検出技術:ShatterSeekアルゴリズムからAI診断まで
数千にも及ぶゲノム断片が入り乱れるクロモスリプシスの全貌を正確に特定し解読するには、単一の古典的な手法ではなく、高解像度の細胞遺伝学的手法と最新のバイオインフォマティクスを組み合わせた統合的アプローチが必須となります。
細胞遺伝学的検出手法の限界と進化
臨床現場で長年用いられてきたGバンド法(核型分析)は、光学顕微鏡の解像度の限界により、クロモスリプシスが引き起こす極めて複雑な再構成を読み解くことは不可能です。そのため、以下のような高度な分子生物学的手法が導入されています。
- ➤アレイCGHおよびSNPアレイ:患者のDNAと参照DNAを比較し、微細な欠失や重複(CNV)をマッピングします。クロモスリプシスの特徴である「コピー数が急速に低下し、ヘテロ接合性の喪失(LOH)を示す領域」を特定する上で有効です。
- ➤FISH(蛍光in situハイブリダイゼーション):全染色体を異なる色で染め分ける技術であり、異常な染色体の構造やダブルマイナス(ecDNA)の存在を単一細胞レベルで直感的に可視化します。
バイオインフォマティクスと「ShatterSeek」アルゴリズム
近年では次世代シーケンシング、特に全ゲノムシークエンス(WGS)の飛躍的発展により、塩基配列レベルの超高解像度で再構成の切断点(ブレイクポイント)を同定することが可能となりました。しかし、膨大なデータからそれが本当にクロモスリプシスであると統計学的に証明するためには、専用のアルゴリズムが不可欠です。
その代表的なグローバルスタンダードとなっているのが、Harvard Medical SchoolのPeter Park研究室によって開発されたRパッケージ「ShatterSeek」です [10]。PCAWGプロジェクトにおいて2,600以上のWGSデータ解析に用いられ、汎がんにおけるクロモスリプシスの全貌解明に大きく貢献しました [10]。
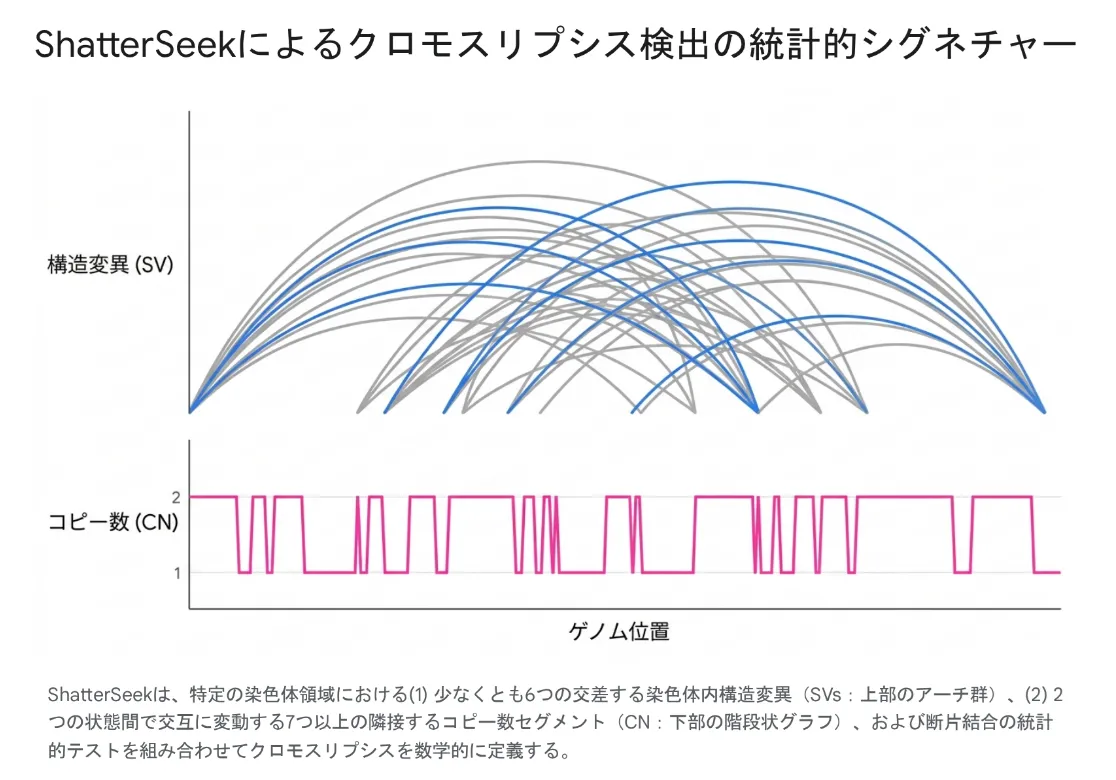
図:ShatterSeekによるクロモスリプシス検出の統計的シグネチャー。構造変異の複雑な交差(上部のアーチ群)と、コピー数の急速な振動(下部の階段状グラフ)を数学的に定義する。
ShatterSeekは、「少なくとも6つの交差する染色体内構造変異」や「2つの状態間で交互に変動する7つ以上の隣接するコピー数セグメント」といった厳密な数学的条件をテストし、他のゲノム異常による擬陽性を排除します [10]。さらに近年では、より安価な全エクソームシーケンス(WES)データからコピー数変異のプロファイルのみを抽出し、AI(ディープラーニング)を用いてクロモスリプシスを予測する「GECNVNet」といった技術も開発され、臨床現場での予後予測モデルとしての実装が進んでいます。
11. 次世代の治療標的としての可能性と未来への展望
クロモスリプシスは、がん細胞に対して無尽蔵の適応力と治療抵抗性を与える強力なドライバーですが、同時にこの極端なゲノム不安定性は、細胞分裂において致命的なエラーを回避し続けなければならないがん細胞自身の「アキレス腱」にもなり得ます。近年の分子メカニズムの深い解明により、特定の分子を直接標的とした新たな治療戦略が現実味を帯びてきています。
N4BP2の阻害を通じたゲノムカオスの予防
第一の有望なアプローチは、ゲノム粉砕の引き金(Spark)となるN4BP2ヌクレアーゼの阻害です。これまでは、既にゲノムが粉砕されてしまった後の「結果」に対処するしかありませんでした。しかし、N4BP2を特異的に阻害する薬理学的なアプローチが確立されれば、がん細胞内で小核が崩壊したとしても、染色体が粉砕されるのを根本から防ぐことができます。これは、ecDNAの爆発的な生成を抑制し、標的治療薬に対する耐性の獲得を未然に防ぐことで、既存のがん治療の効果を長期的に持続させる戦略として極めて有望です。
CIP2A-TOPBP1阻害とDNA修復欠損を突く「合成致死」
第二に、有糸分裂において粉砕された染色体を主核に再統合する足場となるCIP2A-TOPBP1複合体の相互作用阻害です。特に、BRCA1遺伝子やBRCA2遺伝子に変異を持つような相同組換え修復欠損がんに対して、この複合体を阻害する化合物を投与することで、がん細胞特異的に合成致死(Synthetic lethality)を誘導できることが実証されています。
第三に、PARP阻害薬などを利用した合成致死性アプローチも臨床試験等で活発に検証されています。骨肉腫や神経芽腫など、クロモスリプシスが予後を規定する悪性度の高い小児・若年性がんにおいて、これらのDNA修復阻害薬は、極限まで高まったゲノム不安定性を限界点を超えさせ、がん細胞を特異的に死滅させる有効な治療法となる可能性を秘めています。
よくある質問(FAQ)
🏥 遺伝に関するご不安は専門医にご相談ください
ご自身やご家族の遺伝性疾患リスク、あるいは高度な遺伝子検査の結果解釈についてお悩みではありませんか?
ミネルバクリニックでは、臨床遺伝専門医が最新のゲノム医学に基づき、正確で分かりやすい遺伝カウンセリングを提供しています。
参考文献
- [1] Chromothripsis-Explosion in Genetic Science
- [2] CHROMOTHRIPSIS FROM DNA DAMAGE IN MICRONUCLEI
- [3] Chromothripsis and ecDNA initiated by N4BP2 nuclease fragmenting cytoplasm exposed chromosomes
- [4] Cancer cells’ chamber of secrets: the link between micronuclei, chromothripsis and malignancy
- [5] Chromothripsis is a common mechanism driving genomic rearrangements in primary and metastatic colorectal cancer
- [6] TP53 Abnormalities Are Underlying the Poor Outcome Associated with Chromothripsis in Chronic Lymphocytic Leukemia Patients with Complex Karyotype
- [7] Chromothripsis is correlated with reduced cytotoxic immune infiltration and diminished responsiveness to checkpoint blockade immunotherapy
- [8] Chromothripsis in Healthy Individuals Affects Multiple Protein-Coding Genes and Can Result in Severe Congenital Abnormalities in Offspring
- [9] Chromothriptic cure of WHIM syndrome: Implications for bone marrow transplantation
- [10] Comprehensive analysis of chromothripsis in 2,658 human cancers using whole-genome sequencing






