目次

📍 クイックナビゲーション
細胞が、秋に枯れ葉が静かに木から落ちるように、自らの意思で秩序正しく死んでいく——この現象がアポトーシス(apoptosis:プログラム細胞死)です。私たちの体では毎日およそ100億個の細胞がこの仕組みで安全に片づけられ、新しい細胞と入れ替わっています。胎児の指が分かれるのも、免疫が暴走しないのも、そしてがんを未然に防ぐのも、すべてアポトーシスのおかげです。この記事では、その基本から、がん治療や出生前診断(NIPT)への意外なつながりまでを、はじめての方にもわかるように解説します。
Q. アポトーシスとは何ですか?まず結論だけ知りたいです
A. 遺伝子にあらかじめ組み込まれた「秩序ある細胞の自死」のことです。細胞が自分を小さくたたんで片づくため、まわりに炎症を起こしません。発生・免疫・組織の新陳代謝に欠かせず、この仕組みが狂うとがんや神経の病気の原因になります。
- ➤定義と歴史 → 1972年に提唱。「枯れ葉が落ちる」が語源で、能動的でプログラムされた死
- ➤2つの経路 → 外からの命令(外因性)と内側の異常(内因性)。最後はカスパーゼが解体
- ➤ネクローシスとの違い → 炎症を起こすか起こさないかが決定的な差
- ➤がんとの関係 → がんは「死ねなくなった細胞」の病気。治療は死を再起動させる
- ➤NIPTとの関係 → NIPTで読む「胎児DNA」は胎盤のアポトーシスが出した断片そのもの
1. アポトーシスとは:プログラムされた細胞死
アポトーシスとは、細胞が特定のシグナルを受け取って自らを能動的に分解し、まわりに迷惑をかけずに片づく、遺伝子にプログラムされた細胞死のことです。最大の特徴は炎症を起こさないこと。細胞は小さな膜の袋(アポトーシス小体)に分かれ、隣の細胞やマクロファージ(掃除役の免疫細胞)に静かに食べられて消えていきます。
💡 用語解説:プログラム細胞死(プログラムさいぼうし)
「あらかじめ計画された死」という意味です。細胞は壊れて死ぬだけでなく、必要なときに必要な細胞が、遺伝子の指令に従って整然と死ぬ仕組みを持っています。アポトーシスはその代表格で、ほかにも近年はネクロプトーシスやフェロトーシスなど多くの種類が見つかっています。これらをまとめて制御性細胞死と呼びます。
「枯れ葉が落ちる」——語源と歴史
「アポトーシス」という言葉は、ギリシャ語で「枯れ葉や花びらが落ちる」様子を表す言葉に由来します。1972年、KerrとWyllie、Currieの3人が発表した論文で、周囲に炎症を起こさず細胞が静かに自らを解体していく「能動的でプログラムされた過程」として、はじめて明確に定義されました[1]。
この仕組みが遺伝子に厳密に決められていることを証明したのが、線虫(C. elegans)という小さな生き物の研究です。線虫が成体になる過程では合計1090個の細胞がつくられますが、そのうち決まった131個が、個体差なくほぼ同じタイミングで「予定どおり」死にます。死ぬべき細胞が遺伝子レベルで決まっている——この発見は、アポトーシス研究の出発点となりました。
毎日100億個の細胞が静かに死んでいる
ヒトの胎児が育つとき、手の指は最初「水かき」でつながっています。指と指の間の細胞がアポトーシスで消えることで、5本の指が分かれて完成します。脳では正しくつながれなかった神経細胞が、免疫では自分の体を攻撃しかねないリンパ球が、それぞれアポトーシスで取り除かれます。大人になっても、組織を新鮮に保つために毎日およそ100億個の細胞が死に、安全に処理されています。生まれる細胞と死ぬ細胞のバランスこそが、私たちの健康な状態を支えているのです。
2. アポトーシスの仕組み:2つの経路とカスパーゼ
アポトーシスの実行は、カスパーゼという酵素群が連鎖的にスイッチオンされることに完全に依存しています。そのスイッチを入れる入口が2つあります。細胞の外からの命令で始まる外因性経路(デス受容体経路)と、細胞の内側の異常で始まる内因性経路(ミトコンドリア経路)です。下の図は、この2つの経路が最後に1つの「実行部隊」へ合流する全体像を示しています。
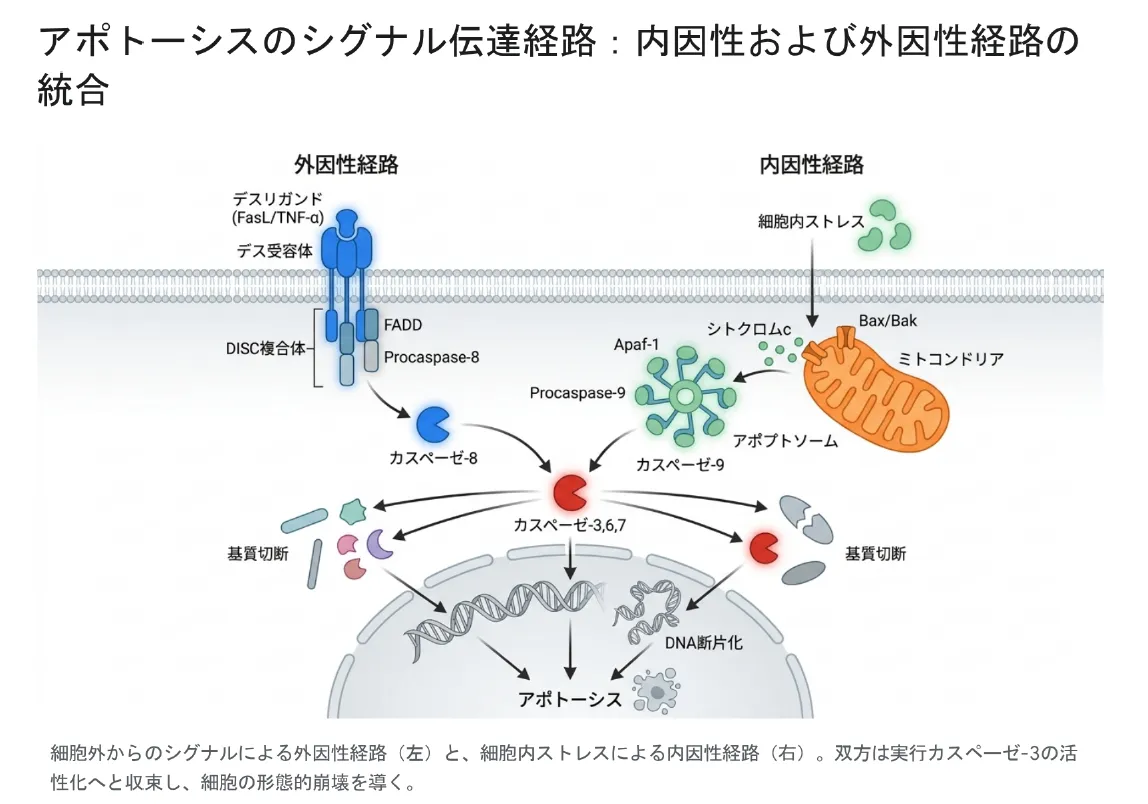
細胞外からの刺激で始まる外因性経路(左)と、細胞内のストレスで始まる内因性経路(右)。どちらも最後は実行カスパーゼ(カスパーゼ-3など)の活性化に合流し、細胞は整然と解体される。
💡 用語解説:カスパーゼ
細胞の中のタンパク質を切る「ハサミ」のような酵素の一群です。ふだんは不活性な状態で待機していますが、アポトーシスの号令がかかると一斉に活性化し、細胞の骨組みやDNAを系統的に切断していきます。入口で号令を出す「開始カスパーゼ(カスパーゼ-8・-9)」と、実際に解体を担う「実行カスパーゼ(カスパーゼ-3・-6・-7)」に分かれています。
外因性経路:外からの「死の命令」を受け取る
外因性経路は、細胞の表面にあるデス受容体(Fas/CD95や腫瘍壊死因子受容体TNFR1など)に、外から「死になさい」というリガンド(FasL・TNF-αなど)が結合することで始まります。結合すると受容体が3つ集まり、内側にFADDなどのアダプターが集合してDISC(死を誘導する信号複合体)という巨大な複合体ができます。ここで開始カスパーゼであるカスパーゼ-8が活性化し、実行カスパーゼに号令を出します。この経路は、免疫細胞が感染した細胞や不要な細胞を排除するときに特に重要です。
内因性経路:ミトコンドリアが「生死のスイッチ」
内因性経路は、DNA損傷・低酸素・強い酸化ストレスなど、細胞内部の深刻な異常で発動します。鍵を握るのはミトコンドリアの外膜です。生死を分けるのがBcl-2ファミリーと呼ばれるタンパク質群のせめぎ合いで、このバランスがミトコンドリアの膜を「閉じておく」か「開ける」かを決めます。
💡 用語解説:Bcl-2ファミリー(生死のシーソー)
細胞の生死を綱引きする3つのグループです。
①生き延びさせる役(抗アポトーシス):Bcl-2、Bcl-xLなど。膜を閉じて死を抑えます。
②膜に穴を開ける役(実行):Bax、Bak。膜に穴を開けて死を進めます。
③センサー役(BH3-only):Bad、Bid、Pumaなど。ストレスを感じて①の働きを止め、②を解き放ちます。
BaxとBakがミトコンドリア外膜に穴を開けると(この現象をMOMPと呼びます)、ふだんは膜の中で電子の運び役をしているシトクロムcが細胞質へ一気に放出されます。シトクロムcはApaf-1というタンパク質と結合し、車輪のような巨大な構造体アポトソーム(死の車輪)をつくります。
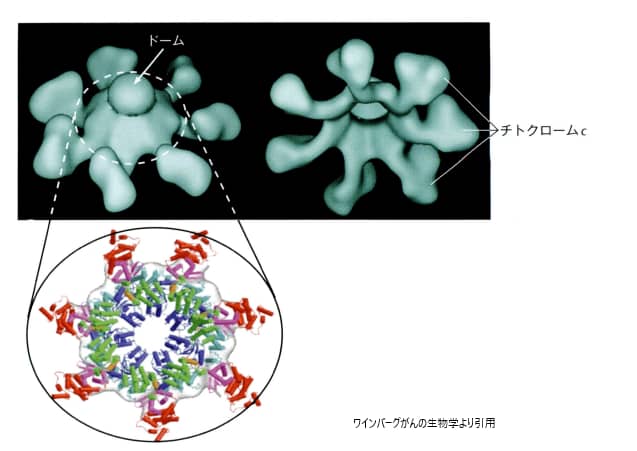
シトクロムcとApaf-1が組み上がってできる7本スポークの車輪状構造「アポトソーム」。中心に開始カスパーゼ-9が呼び込まれ、活性化される。
この車輪の中心に開始カスパーゼカスパーゼ-9が呼び込まれて活性化し、実行カスパーゼへとバトンを渡します。さらにミトコンドリアからはSMAC/DIABLOという分子も放出されます。これはカスパーゼにブレーキをかけている「アポトーシス阻害タンパク質(IAP)」を無力化する役で、いわば死のブレーキを外す働きをします。
実行フェーズ:カスパーゼ-3による「解体作業」
2つの経路はどちらも最後に実行カスパーゼ(特にカスパーゼ-3)を活性化させ、ここで合流します。カスパーゼ-3は細胞解体の「処刑人」として、DNA修復酵素PARPを切ってDNA修復を止め、核膜を支えるラミンを切って核を崩壊させ、さらにDNA分解酵素のストッパー(ICAD)を切ってDNA切断を開始させます。こうして細胞は、後戻りできない形で整然と分解されていきます。なお免疫の現場では、これら2経路に加えて、NK細胞やキラーT細胞がグランザイムBを標的細胞に注入して直接カスパーゼを起動する「パーフォリン/グランザイム経路」も働いています。
3. 細胞に起こる「目に見える変化」
顕微鏡でアポトーシス中の細胞を見ると、はっきりした特徴が順番に現れます。まず細胞全体が縮み(細胞収縮)、表面に泡のような突起が出ます(ブレブ形成:blebbing)。核の中ではDNAがぎゅっと凝集し(ピクノーシス)、やがて核そのものが断片化します(カリオレキシス)。
このとき、カスパーゼに起こされたDNA分解酵素が、DNAを決まった位置で切るため、DNAは約180〜200塩基対の倍数で規則正しく切断されます。これを電気泳動にかけると「はしご(ラダー)」状の模様になるのが、アポトーシスの生化学的な目印です。
💡 用語解説:「Eat-me(私を食べて)」シグナル
アポトーシスの最終段階で細胞は小さな袋(アポトーシス小体)に分かれます。その表面には、ふだんは細胞膜の内側に隠れているホスファチジルセリンという脂質が顔を出します。これが「私を食べてください」という目印になり、マクロファージが中身を漏らさないうちに素早く食べてくれます。だからアポトーシスでは炎症がまったく起こらないのです。
4. ネクローシスなど、他の細胞死との違い
長い間、細胞死は「秩序あるアポトーシス」と「事故的なネクローシス」の2つで理解されてきました。しかし近年、両者の中間のようなプログラムされたネクローシスも次々と見つかっています。最大の見分け方は、炎症を起こすかどうかです。
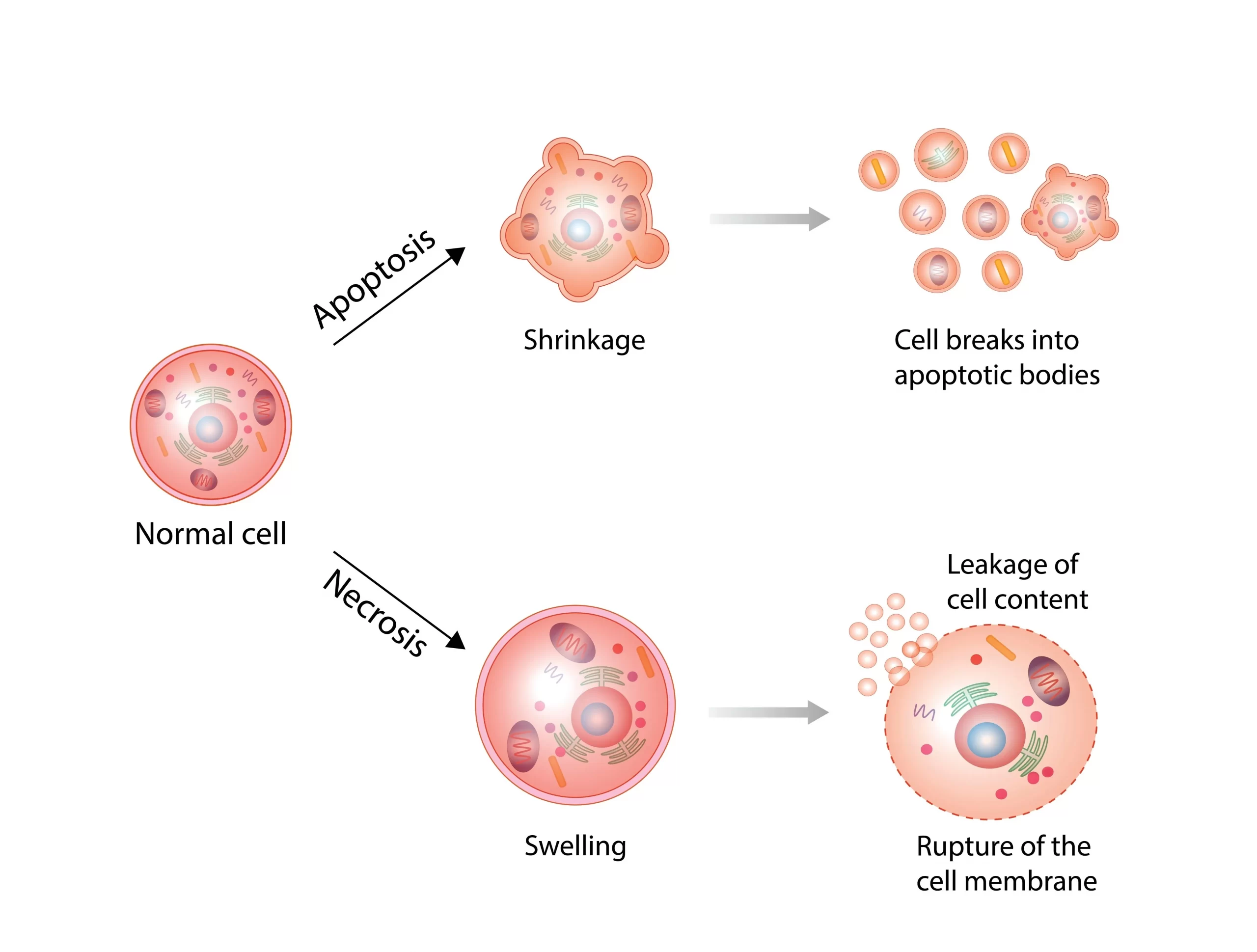
アポトーシスは細胞が縮んで小体に分かれ静かに片づくのに対し、ネクローシスは膨張して破裂し、中身が漏れて強い炎症を起こす。
💡 用語解説:ネクローシスとDAMPs
ネクローシス(壊死)は、強い熱・虚血・物理的損傷などで細胞が事故的に死ぬことです。膜が破れて中身が漏れ出し、その内容物がDAMPs(ダメージ関連分子パターン)として周囲に「警報」を発し、強い炎症と二次的な組織損傷を引き起こします。アポトーシスとは正反対に、まわりを巻き込む死だといえます。
| 特徴 | アポトーシス | ネクローシス | ネクロプトーシス | パラプトーシス |
|---|---|---|---|---|
| 形の変化 | 縮小・ブレブ形成 | 膨張・破裂 | 膨張・破裂 | 細胞質に大きな空胞 |
| 細胞膜 | 保たれる(小体形成) | 早期に破綻 | 早期に破綻 | 後期まで保たれる |
| カスパーゼ | 必須 | 不要 | 不要(むしろ抑制時) | 不要 |
| 炎症 | 起こさない | 強い | 強い | 通常なし |
| 主な分子 | カスパーゼ・Bcl-2 | ATP枯渇・物理損傷 | RIPK1・RIPK3・MLKL | IGF-1R・HMGB1放出 |
アポトーシスとネクローシスは完全に独立しているわけではありません。アポトーシスが進行中でも、細胞のエネルギー(ATP)が尽きると最後までやり遂げられず、ネクローシスへ移行することがあります(アポトーシス・ネクローシス連続体)。また、アポトーシスで死んだ細胞がすぐに食べられないと、最後に膜が破れて「二次性ネクローシス」になることも知られています。これら多彩な細胞死は、それぞれ別の記事で詳しく解説しています。
5. がんとアポトーシス:「死ねなくなった細胞」の病気
がんは「異常に増える」だけの病気だと思われがちですが、もう一つの本質は「死ねなくなること(アポトーシス回避)」にあります。本来、DNAに深刻な傷を負った細胞は、自らアポトーシスで死んでがん化を防ぎます。その安全装置を壊した細胞が、際限なく増え続けてがんになるのです。
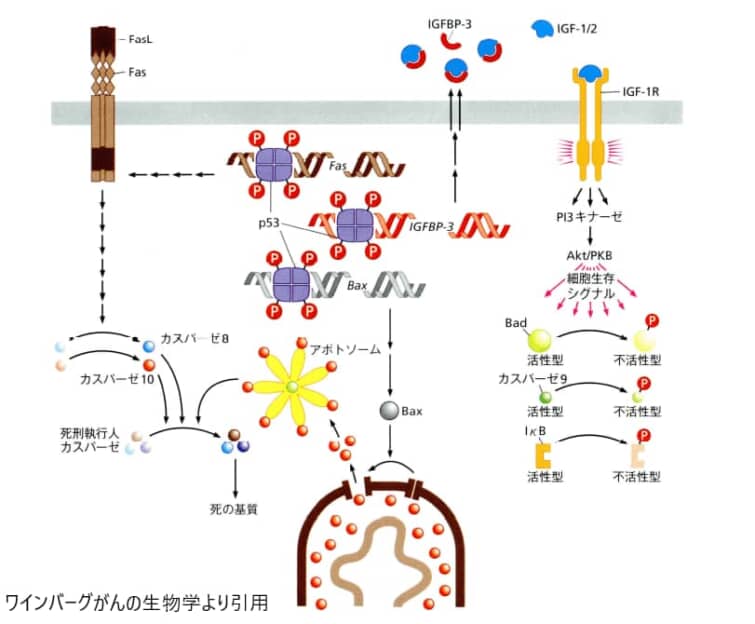
「ゲノムの守護神」p53は、複数の経路を通じてアポトーシスを起動し、傷ついた細胞ががん化する前に排除する。
💡 用語解説:p53(がん抑制遺伝子の代表)
p53は「ゲノムの守護神」とも呼ばれるタンパク質で、設計図であるDNAに傷がつくと活性化します。修復できそうなら細胞の増殖を一時停止して直し、修復が無理ならアポトーシスを起こして細胞を始末します。ところが、すべてのヒトのがんの約50%でこのp53(遺伝子名TP53)が変異して機能を失っていることが知られています[4]。
💡 用語解説:ミスセンス変異
DNAの文字(塩基)が1つ書き換わることで、できあがるタンパク質のアミノ酸が1個だけ別の種類に置き換わってしまうタイプの遺伝子変異です。たった1文字の違いでも、タンパク質の形がゆがんで本来の働きを失うことがあります。p53の機能を奪う変異の多くは、このミスセンス変異です。
もう一つのよくある回避手段が、生き延びさせる役のタンパク質Bcl-2の過剰発現(アップレギュレーション)です。Bcl-2が増えすぎると、死の引き金役であるBaxやBakが押さえ込まれ続け、ミトコンドリアの穴が開かず、細胞は死を免れて増殖を続けます。なお、ひとくちに「がん」と言っても上皮性の「癌」と血液がんなどでは性質が異なります(「がん」と「癌」の違いを参照)。
6. アポトーシスを利用した、がん治療の最前線
がん細胞が止めてしまった「死のプログラム」を、もう一度起動させる——これが現代のがん分子標的治療で最も注目される戦略の一つです。主なアプローチを紹介します。
① p53の機能を取り戻す
失われたp53を補う遺伝子治療として、正常なp53遺伝子をウイルスで運び込む薬(Gendicineなど)が中国で承認・使用されています。また、p53を持たないがん細胞だけで増えて細胞を壊すよう改変したウイルス(ONYX-015)や、p53を分解させてしまうMDM2の働きを止める小分子薬(Nutlins)の開発も進んでいます。
② Bcl-2を直接ブロックする——ベネトクラクス
近年もっとも鮮やかな成功例が、Bcl-2を狙い撃ちするベネトクラクス(Venetoclax)です。これはセンサー役(BH3-only)の形をまねた「BH3ミメティック」で、過剰なBcl-2に強く結合し、それまで拘束されていたBaxやBakを一気に解き放ちます。するとがん細胞自身に備わった死のプログラムが強制的に起動します。慢性リンパ性白血病(CLL)や急性骨髄性白血病(AML)で高い効果を上げています[5]。がん細胞が頼っていた「生き延びるための盾」を逆手に取る、エレガントな治療法です。
💡 用語解説:BH3ミメティック
「BH3-onlyタンパク質(死を促すセンサー役)のまねをする薬」という意味です。本物のセンサーそっくりの形をしているため、Bcl-2の結合ポケットにすっぽりはまり込み、Bcl-2が握りしめていた「死の実行役」を解放します。狙った分子だけに効くよう設計された分子標的薬の一種です。

7. アポトーシスとNIPT(新型出生前診断)の深い関係
ここまで細胞生物学とがんの話をしてきましたが、このミクロの「細胞死」は、当院が専門とするNIPT(新型出生前診断)の土台にもなっています。意外に思われるかもしれませんが、NIPTで読み取る「胎児DNA」の正体は、胎盤の細胞がアポトーシスを起こして放出したDNAの断片そのものなのです。
💡 用語解説:セルフリー胎児DNA(cffDNA)
妊婦さんの血液(血漿)の中を漂っている、胎盤由来のごく短いDNA断片のことです。1997年にDennis Loらが発見しました。母体から採血するだけで解析できるため、ダウン症候群などの染色体の数の変化を、おなかに針を刺さずに高い精度で調べられます。この断片は約143〜166塩基対と非常に短いのが特徴で、これはアポトーシスの規則正しいDNA切断を経て生じた証拠です。
「胎児DNA」が母体血に届くまで
なぜ「偽陽性」「偽陰性」が起こるのか
このメカニズムを理解すると、NIPTの限界も論理的に見えてきます。NIPTが調べているのは厳密には「胎児の細胞」ではなく「胎盤の細胞がアポトーシスで放出したDNA」です。ところが胎児本体と胎盤は、発生のごく初期に分かれるため、まれに胎盤だけが染色体異常を持ち、胎児本体は正常という状態が起こります。
💡 用語解説:限局性胎盤モザイク(CPM)
染色体異常が胎盤の細胞にだけ限られ、胎児本体には及んでいない状態です。全妊娠の約1〜2%で起こるとされます。胎盤の異常な細胞がアポトーシスでDNAを出すと、NIPTは「陽性」と判定しますが、赤ちゃん自身は健康——という偽陽性の主な原因がこれです。だからNIPTは「確定診断」ではなく「スクリーニング(ふるい分け)」なのです。
NIPTの判定と確定診断が食い違う「不一致」は全症例の約0.1%で起こりますが、その最大の理由がこの胎児と胎盤のズレです。検査の精度を高める解析手法(COATE法など)や、胎児フラクション(cffDNAの割合)が足りないときの判定保留の運用は、いずれもこの生物学的事実をふまえた仕組みです。
最先端:妊娠高血圧腎症の早期予測へ
アポトーシス由来のcffDNAの解析は、赤ちゃんの染色体だけでなく、お母さんの健康を守る方向にも広がっています。胎盤の形成がうまくいかず慢性的な酸素不足に陥ると、胎盤の細胞が異常なアポトーシス/ネクローシスを起こし、血液中のDNAの「出どころの比率」が変化します[7]。この変化をとらえることで、重い妊娠合併症である妊娠高血圧腎症を、症状が出る数か月前に予測できる可能性が報告されています。
妊娠初期からの早期NIPTの実現も含め、ミクロの細胞死を読み解く技術が、周産期医療を大きく前進させようとしています。検査をブラックボックスとして扱うのではなく、なぜ偽陽性が起こるのか、判定保留が何を意味するのかまで理解したうえで、ご家族と一緒に考えること——それが私たちの遺伝カウンセリングの役割だと考えています。
8. よくある誤解
誤解①「アポトーシス=悪いこと」
細胞が死ぬと聞くと悪いことに思えますが、アポトーシスは健康を保つために不可欠な仕組みです。むしろ起こらなさすぎる(がん)ことも、起こりすぎる(神経変性)ことも問題になります。
誤解②「がんは増えるだけの病気」
増殖の異常だけでなく、「死ねなくなる(アポトーシス回避)」こともがんの本質です。だからこそ「死を再起動させる」治療が成り立ちます。
誤解③「ネクローシスと同じ細胞死」
同じ「死」でも全く別物です。炎症を起こすか・起こさないかが決定的に違い、治療への意味づけも正反対になります。
誤解④「NIPTは胎児の細胞を直接見ている」
実際に見ているのは胎盤のアポトーシス由来のDNA断片です。この事実が、偽陽性が起こる理由や、確定診断が別に必要な理由を説明します。
9. 臨床遺伝専門医からのメッセージ
よくある質問(FAQ)
🏥 出生前診断・遺伝のご相談
アポトーシスを土台とするNIPTをはじめ、出生前診断や遺伝に関するご相談は、
臨床遺伝専門医が在籍するミネルバクリニックへお気軽にどうぞ。
参考文献
- [1] Elmore S. Apoptosis: A Review of Programmed Cell Death. Toxicol Pathol. 2007;35(4):495-516. [PMC2117903]
- [2] Apoptosis: A Comprehensive Overview of Signaling Pathways, Morphological Changes, and Physiological Significance and Therapeutic Implications. PMC. 2024. [PMC11592877]
- [3] Intrinsic and Extrinsic Pathways of Apoptosis. Thermo Fisher Scientific. [Thermo Fisher]
- [4] Wang Z, Sun Y. Targeting p53 for Novel Anticancer Therapy. Transl Oncol. 2010;3(1):1-12. [PMC2822448]
- [5] Lasica M, Anderson MA. Review of Venetoclax in CLL, AML and Multiple Myeloma. J Pers Med. 2021;11(6):463. [PMC8225137]
- [6] Cell-free fetal DNA in the maternal circulation originates from the cytotrophoblast. PMC. [PMC4498868]
- [7] Can Cell-free Fetal DNA Predict Preeclampsia Before Clinical Onset? Preeclampsia Foundation (Nature Medicine, 2025). [Preeclampsia Foundation]
- [8] What is apoptosis, and why is it important? PMC. [PMC1120576]






