目次
- 1 1. ヒスチジンメチル化とは?──半世紀ぶりに解けはじめた「謎」
- 2 2. 担い手は4つの酵素──2つのファミリーと役割分担
- 3 3. SETD3──アクチン・平滑筋収縮・原発性難産・ウイルス感染のハブ
- 4 4. METTL9──亜鉛のバランスとがんを操る「広く効く」酵素
- 5 5. METTL18──翻訳の「あえてのブレーキ」でタンパク質の品質を守る
- 6 6. CARNMT1──RNA代謝を根っこから支える修飾酵素
- 7 7. 「消す酵素」はあるのか──脱メチル化をめぐる議論
- 8 8. どうやって測るのか──検出を阻んだ「技術の壁」と突破口
- 9 9. 遺伝医療との接点──いまは「研究段階」だからこそ知っておきたいこと
- 10 10. よくある誤解
- 11 よくある質問(FAQ)
- 12 参考文献
- 13 関連記事

📍 クイックナビゲーション
タンパク質のヒスチジンメチル化は、長年「あるのは知っているのに、それを行う酵素の正体がわからない」という状態が半世紀ほど続いた、翻訳後修飾(タンパク質が作られた後に受ける化学的な飾りつけ)の一つです。近年になってSETD3・METTL9・METTL18・CARNMT1という4つの主要な酵素が相次いで見つかり、この修飾が細胞骨格・翻訳・亜鉛のバランス・RNA代謝といった細胞の根幹に関わっていること、そしてがん・原発性難産・ウイルス感染など幅広い病態と結びつくことがわかってきました。この記事では、その全体像を遺伝専門医の視点でやさしく整理します。
Q. ヒスチジンメチル化とは何ですか?まず結論だけ知りたいです
A. タンパク質を構成するアミノ酸「ヒスチジン」に、メチル基(CH₃)という小さな部品を1つ付け加える化学修飾です。付ける位置によって「1-メチルヒスチジン」と「3-メチルヒスチジン」の2種類ができ、これによってタンパク質の電気的な性質や亜鉛などの金属を捕まえる力が変わります。SETD3・METTL9・METTL18・CARNMT1という酵素が担い、細胞骨格・翻訳・免疫・RNA代謝の調節に関わります。
- ➤修飾の正体 → ヒスチジンの環にある2つの窒素(N1=π位/N3=τ位)のどちらかにメチル基が付く
- ➤担い手は4酵素 → METTL9・CARNMT1がN1位、SETD3・METTL18がN3位を担当
- ➤病気とのつながり → がんの進行、オキシトシンが効きにくい原発性難産、エンテロウイルス感染などに関与
- ➤意外な広がり → ヒトの全メチル化イベントの約13%を占め、決して珍しい修飾ではない
- ➤残る課題 → 「消す酵素(脱メチル化酵素)」の正体や、専用の検出ツールの開発は研究途上
1. ヒスチジンメチル化とは?──半世紀ぶりに解けはじめた「謎」
私たちの体を作るタンパク質は、20種類のアミノ酸が数珠つなぎになってできています。しかしタンパク質は作られたそのままの姿で働くわけではなく、あとから小さな部品を付けたり外したりして「機能を微調整」されます。これを翻訳後修飾と呼びます。その代表がメチル基(CH₃)を付ける「メチル化」で、これまで研究の中心はリジンとアルギニンという2つのアミノ酸でした。ところが近年、3番目のアミノ酸ヒスチジンへのメチル化が、想像以上に広く細胞を制御していることがわかってきました。実際、ヒトの細胞で起こる全メチル化イベントのうちおよそ13%がヒスチジンの修飾だと見積もられています[3]。
ヒスチジンメチル化そのものは、決して新しい発見ではありません。1967年には、さまざまな脊椎動物の筋肉タンパク質であるアクチンとミオシンから、天然の成分としてメチル化されたヒスチジン(3-メチルヒスチジン)が見つかっていました[2]。しかし、この修飾に生物学的にどんな意味があるのか、そして「誰が(どの酵素が)」付けているのかは、その後50年以上も大きな謎のまま残されていました[2]。担い手の酵素が見つかりはじめたのは、ようやく2010年代以降のことです。
💡 用語解説:翻訳後修飾(ほんやくごしゅうしょく/PTM)
設計図(遺伝子)をもとにタンパク質が組み立てられる工程を「翻訳」と呼びます。翻訳後修飾とは、その完成品にあとから化学的な「飾りつけ」を加えて働きを調整する仕組みのことです。メチル化・リン酸化・ユビキチン化などがあり、スイッチのオン/オフ、他のタンパク質との結合の強さ、分解のタイミングなどを細かくコントロールします。1つの遺伝子から、状況に応じて何通りもの「働き方」を引き出すための、細胞の重要な調節手段です。
2つの窒素、2種類のメチルヒスチジン
ヒスチジンの側鎖には「イミダゾール環」という五角形の構造があり、そこに窒素原子が2つあります。メチル基はこのどちらかの窒素に付き、付く場所によって別の分子ができます。側鎖に近い側の窒素(N1位、別名π位)に付いたものが「1-メチルヒスチジン」、遠い側の窒素(N3位、別名τ位)に付いたものが「3-メチルヒスチジン」です[3]。たった1個のメチル基でも、イミダゾール環の疎水性やプロトン化の状態(電荷)が変わり、さらに亜鉛など2価金属イオンへの結合力が大きく変化するため、生化学的な影響は決して小さくありません[18]。
%3C/text%3E%3Cline x1='400' y1='118' x2='400' y2='146' stroke='%237f8c8d' stroke-width='2'/%3E%3Cpolygon points='393,144 407,144 400,154' fill='%237f8c8d'/%3E%3Crect x='320' y='150' width='160' height='58' rx='10' fill='%2334495e'/%3E%3Ctext x='400' y='177' text-anchor='middle' font-size='16' fill='%23ffffff' font-weight='bold'%3Eヒスチジン (His)%3C/text%3E%3Ctext x='400' y='197' text-anchor='middle' font-size='11' fill='%23ecf0f1'%3Eイミダゾール環に N1(π)・N3(τ)%3C/text%3E%3Cline x1='340' y1='210' x2='232' y2='250' stroke='%232980b9' stroke-width='2'/%3E%3Cpolygon points='228,242 242,247 230,258' fill='%232980b9'/%3E%3Cline x1='460' y1='210' x2='568' y2='250' stroke='%231e8449' stroke-width='2'/%3E%3Cpolygon points='572,242 558,247 570,258' fill='%231e8449'/%3E%3Crect x='70' y='258' width='300' height='170' rx='12' fill='%23eaf4fc' stroke='%233498db' stroke-width='3'/%3E%3Ctext x='220' y='288' text-anchor='middle' font-size='16' fill='%232980b9' font-weight='bold'%3EN1(π)位のメチル化%3C/text%3E%3Ctext x='220' y='316' text-anchor='middle' font-size='15' fill='%232c3e50' font-weight='bold'%3EMETTL9 / CARNMT1%3C/text%3E%3Ctext x='220' y='344' text-anchor='middle' font-size='14' fill='%2334495e'%3E→ 1-メチルヒスチジン%3C/text%3E%3Ctext x='220' y='378' text-anchor='middle' font-size='12' fill='%235b6b7b'%3E亜鉛の結合を調節/RNA代謝を制御%3C/text%3E%3Ctext x='220' y='402' text-anchor='middle' font-size='12' fill='%235b6b7b'%3E(S100A9・SLC39A7・U2AF1 など)%3C/text%3E%3Crect x='430' y='258' width='300' height='170' rx='12' fill='%23e8f8f0' stroke='%2327ae60' stroke-width='3'/%3E%3Ctext x='580' y='288' text-anchor='middle' font-size='16' fill='%231e8449' font-weight='bold'%3EN3(τ)位のメチル化%3C/text%3E%3Ctext x='580' y='316' text-anchor='middle' font-size='15' fill='%232c3e50' font-weight='bold'%3ESETD3 / METTL18%3C/text%3E%3Ctext x='580' y='344' text-anchor='middle' font-size='14' fill='%2334495e'%3E→ 3-メチルヒスチジン%3C/text%3E%3Ctext x='580' y='378' text-anchor='middle' font-size='12' fill='%235b6b7b'%3E細胞骨格・翻訳マシンを制御%3C/text%3E%3Ctext x='580' y='402' text-anchor='middle' font-size='12' fill='%235b6b7b'%3E(アクチン His73・リボソーム RPL3)%3C/text%3E%3C/svg%3E)
メチル基の供与体SAMを使い、N1(π)位はMETTL9・CARNMT1が、N3(τ)位はSETD3・METTL18が担当します。同じ「ヒスチジンメチル化」でも、位置が違えば別の分子・別の生理機能になります。
💡 用語解説:1-メチルと3-メチルの「番号のややこしさ」
この分野には昔からの落とし穴があります。生化学・栄養学の分野と、有機化学(IUPAC)の分野とで、イミダゾール環の窒素の番号の付け方が逆になっているため、文献によって「1-メチル」と「3-メチル」の呼び名がπ位・τ位と入れ替わって見えることがあります。本記事はヒスチジンメチル基転移酵素の研究分野で標準的な表記に統一し、N1=π位=1-メチルヒスチジン(METTL9・CARNMT1が担当)、N3=τ位=3-メチルヒスチジン(SETD3・METTL18が担当)として説明します。
2. 担い手は4つの酵素──2つのファミリーと役割分担
ヒスチジンメチル化を担う酵素は、構造と進化の由来から大きく2つのファミリーに分かれます。1つは「SETドメイン」を持つグループ、もう1つは「7BS(7本鎖βシート)メチル基転移酵素」と呼ばれるグループです[4]。どちらも共通の「メチル基の供与体」であるSAM(S-アデノシルメチオニン)から、メチル基を1つ取り出してヒスチジンに渡します。おもしろいのは、この4つの酵素がそれぞれ担当する窒素(N1位かN3位か)をきっちり区別している点です。METTL9とCARNMT1はN1(π)位を、SETD3とMETTL18はN3(τ)位を修飾します[3]。
SETドメインを持つ酵素はヒトに約55種類あり、その大半はヒストンのリジンを修飾する酵素(ヒストンリジンメチル基転移酵素)です[4]。そのなかで、ヒトで初めて「ヒスチジン」を修飾する酵素として同定されたSETD3は、いわば例外的な存在です。活性中心のわずかな構造の違いによって、リジンではなくヒスチジンを狙い撃ちできるようになりました[5]。一方の7BSファミリー(METTL9・METTL18・CARNMT1)は、ねじれた7本のβシートという独特の骨格を共有し、酵素ごとに違う「標的配列の目印(モチーフ)」を読み取って基質を選びます[4]。
💡 用語解説:SAM(S-アデノシルメチオニン)
細胞の中で「メチル基を配る宅配便」の役割をする分子です。メチル基転移酵素は、このSAMからメチル基を1つ受け取り、DNA・RNA・タンパク質などの決められた場所に渡します。メチル基を渡し終えたSAMはSAH(S-アデノシルホモシステイン)に変わります。ヒスチジンメチル化を含むほとんどのメチル化反応が、このSAMを共通の材料として使っています。
3. SETD3──アクチン・平滑筋収縮・原発性難産・ウイルス感染のハブ
🔍 関連記事:SETD3遺伝子/ACTB遺伝子(βアクチン)/アクチンとは/アクチノパチー
SETD3は、細胞の形と動きを支える骨組みアクチンの、73番目のヒスチジン(His73)をN3(τ)位でメチル化する酵素です[7]。じつは初期の研究(2011年)ではSETD3はヒストンのリジンを修飾する酵素だと誤って報告されていましたが、その後これはデータの読み違いだったことが証明され、本当の生理的な基質はアクチンであることが確定しています[5]。この「リジンかヒスチジンか」の切り替えは、活性中心のごくわずかな構造の違いに支えられており、実際に活性中心の2アミノ酸(N256とW274)を人工的に置き換えると、酵素の好みがヒスチジンからリジンへと約18,000倍も入れ替わることが示されました[6]。
SETD3がアクチンと結合すると、アクチンの一部が本来の折りたたみをほどいてまっすぐ引き伸ばされた形に変わり、His73が酵素のポケットの奥に正確にはまり込みます[7]。このメチル化はアクチン単量体のヌクレオチド交換をゆっくりにし、アクチンが繊維へと重合していく過程を穏やかに整えます[5]。つまりヒスチジンメチル化が、細胞骨格という「建物の鉄骨」の組み立て速度を微調整しているのです。
平滑筋の収縮と「原発性難産」
このアクチンのメチル化が最もはっきりと形になるのが、筋肉の収縮です。SETD3遺伝子を失わせた雌マウスでは、複数の組織でアクチンのHis73メチル化が完全に消え、重い原発性難産を起こしました[8]。正常なマウスが交尾後19日で無事に出産するのに対し、SETD3を欠くマウスは20日を過ぎても胎児を子宮内に残したまま出産が完了せず、しかもオキシトシンなどの陣痛促進薬にも反応しにくいのが特徴でした[8]。ヒトの子宮平滑筋細胞を使った実験でも、SETD3を減らすと収縮が著しく妨げられたことから、ヒスチジンメチル化が平滑筋のはたらきに欠かせないことが示されています[8]。
💡 用語解説:原発性難産(げんぱつせいなんざん)
子宮の筋肉(平滑筋)が十分に収縮せず、お産がうまく進まない状態を指します。ここで紹介したのはマウスとヒト細胞を用いた研究段階の知見であり、ヒトの難産すべてがこの仕組みで説明できるわけではありません。ただし「アクチンのヒスチジンメチル化が筋収縮に必要」という発見は、これまで原因のはっきりしなかった分娩の異常を、分子レベルで理解するための新しい手がかりを与えています。

エンテロウイルス感染に絶対に必要な「宿主因子」
SETD3のもう一つの顔は、ウイルスの共犯者としての役割です。SETD3は、ポリオウイルス・コクサッキーウイルスB3・エンテロウイルスA71などが増えるために欠かせない「宿主因子」であり、SETD3を欠くマウスはこれらのウイルス感染に対して完全な抵抗性を示しました[9]。仕組みとしては、SETD3がウイルスの2Aプロテアーゼというタンパク質分解酵素と直接がっちり結合します。興味深いことに、この結合はSETD3のメチル化を行う活性とは無関係で、複合体そのものが物理的にウイルスの増殖を後押ししていると考えられています[9]。
この発見は創薬の面でも重要です。SETD3の酵素活性をつぶすのではなく、SETD3とウイルスタンパク質の結合面(タンパク質間相互作用)だけを邪魔する化合物は、ウイルス自身の遺伝子ではなく宿主の分子を狙うため、ウイルスが変異で薬に耐性を持ちにくい「宿主指向型の抗ウイルス薬」の理想的なモデルになり得ると期待されています[9]。
4. METTL9──亜鉛のバランスとがんを操る「広く効く」酵素
🔍 関連記事:METTL9遺伝子と酵素/フェロトーシスとは/アポトーシス
SETD3がアクチン専用だったのに対し、METTL9はとても幅広い相手を修飾する酵素で、ヒトのタンパク質で起こるN1(π)位ヒスチジンメチル化の大半を担っています[3]。METTL9は「xHxH(xは小さな側鎖のアミノ酸、Hはヒスチジン)」という特定の並びを目印にして、2番目のヒスチジンをメチル化します[3]。この「xHxH」という配列は、生き物の世界で亜鉛イオンを捕まえる場所としてよく使われるため、METTL9によるメチル化は「亜鉛を捕まえる力をわざと弱める」という意味を持ちます[10]。
代表的な相手が、免疫や抗菌に関わるS100A9というタンパク質です。S100A9のHis107がメチル化されても、パートナー分子S100A8と組む能力は保たれますが、亜鉛への結合力は大きく下がります[10]。これによって、その場所にある「遊離の亜鉛」の量や、亜鉛を奪って細菌をやっつける働きが動的に調整されると考えられています。
亜鉛の輸送体を書き換えて「細胞死に強くする」
もう一つの重要な相手が、亜鉛を運ぶトランスポーターSLC39A7(ZIP7)です。METTL9がSLC39A7のヒスチジンをメチル化すると、細胞内の亜鉛の流れが変わり、亜鉛のバランス全体が組み替えられます[12]。この変化はPERK/ATF4という小胞体ストレスの経路を動かし、その先でシスチンの取り込みを増やしてグルタチオン(細胞を守る抗酸化物質)を作らせ、結果として細胞はフェロトーシスという鉄依存性の細胞死に強くなるのです[12]。
💡 用語解説:フェロトーシス
鉄(Fe)に依存して起こる、比較的新しく発見された細胞死のかたちです。細胞膜の脂質が酸化されて壊れることで死に至ります。がん細胞にとっては「弱点」にもなり得るため、あえてフェロトーシスを起こして治療する研究が進んでいます。逆に言えば、METTL9のようにフェロトーシスへの耐性を与える仕組みは、がん細胞を生き延びさせる方向にはたらくことになります。
この「細胞死に強くする」性質は、がんにとって好都合です。実際、METTL9は有望ながん原遺伝子(オンコジーン)として働くことが複数の研究で示され、膵臓がん・肝臓がん・胃がんではMETTL9の発現が高い患者ほど予後が悪い傾向が報告されています[11]。とりわけ硬性胃がん(スキルス胃がん)では、転移した細胞でMETTL9が主にミトコンドリアに集まり、腹膜への播種と強く相関していました[13]。マウスの移植腫瘍でMETTL9を欠かせると腫瘍の増殖が著しく抑えられ、強い抗腫瘍免疫が引き出されたことから、METTL9の酵素活性を狙う小分子は、新しい作用の抗がん・抗転移薬になり得ると考えられています[11]。
5. METTL18──翻訳の「あえてのブレーキ」でタンパク質の品質を守る
🔍 関連記事:ユビキチン‐プロテアソーム系/リン酸化/PI3K-AKT-mTOR経路
METTL18は、タンパク質を作る工場「リボソーム」の部品であるRPL3というタンパク質のHis245をN3(τ)位でメチル化します[15]。この仕組みは酵母(パン酵母)のHpm1という酵素の研究から連綿と続くもので、2010年にHpm1が酵母のRpl3のHis243を修飾する酵素として同定されたのが、世界で最初に見つかったタンパク質ヒスチジンメチル基転移酵素でした[14]。ヒトのMETTL18は、この機能をさらに洗練させ、リボソームができあがる途中の中間体に結合して修飾を行い、最適なリボソーム生合成を保証します[15]。
このメチル化の効果は、とても具体的です。修飾されたリボソームは、チロシン(Tyr)のコドンを通過するときだけ、あえて翻訳の速度を落とすようになります[14]。一見すると効率が悪くなるように思えますが、これは重要な「翻訳品質管理」の仕組みです。リボソームが一瞬立ち止まることで、作られたばかりのタンパク質の鎖が、リボソームの出口から出ながら正しい立体構造に折りたたまれるための十分な時間が確保されるのです[14]。
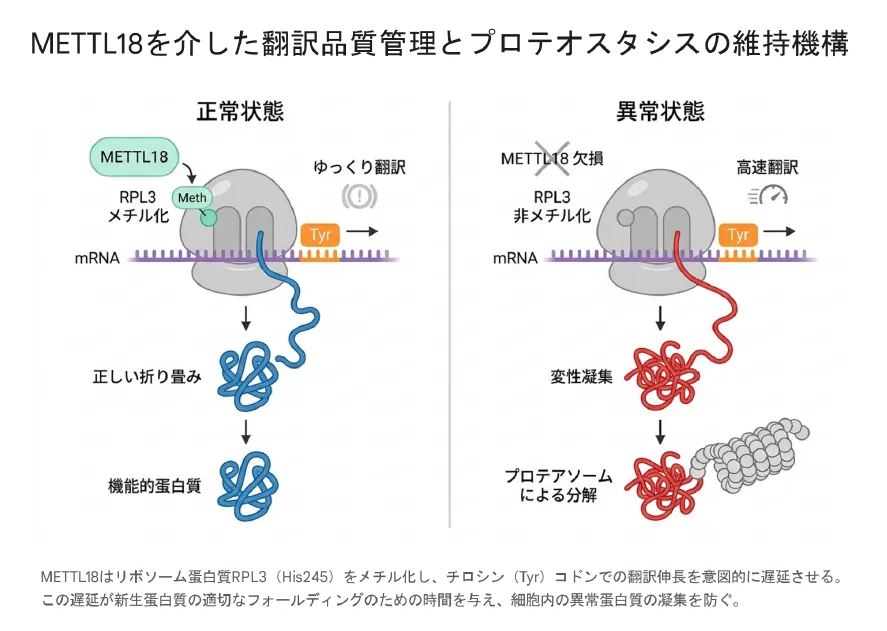
METTL18はリボソームタンパク質RPL3(His245)をメチル化し、チロシン(Tyr)コドンでの翻訳伸長を意図的に遅らせます。この遅延が新生タンパク質の折りたたみに必要な時間を与え、細胞内の異常タンパク質の凝集を防ぎます。
実際、METTL18を失った細胞ではリボソームがTyrコドンを減速せずに走り抜けてしまうため、チロシンを多く含むタンパク質の折りたたみが追いつかず、細胞全体のタンパク質の健全さ(プロテオスタシス)が損なわれます[14]。折りたたみに失敗したタンパク質は異常な塊を作り、細胞はこれをユビキチン‐プロテアソーム系という「ゴミ処理装置」で積極的に分解して対処します[14]。
HER2陰性乳がんの転移との関わり
このプロテオスタシス制御は、がんの悪性化とも結びつきます。乳がんの遺伝子発現を解析した研究では、METTL18がHER2陰性乳がんで特異的に増幅しており、発現の上昇が予後不良と強く結びついていました[16]。その転移促進の仕組みは、METTL18によるRPL3のメチル化がシャペロンタンパク質HSP90を高く保ち、それを介してアクチンの重合が促され、さらにHSP90とがん遺伝子産物Srcキナーゼが足場の上で相互作用してSrcが活性化される、という一連のネットワーク(METTL18‑HSP90‑アクチン‑Src軸)に依存しています[16]。活性化したSrcは下流のPI3KやSTAT3の経路を動かし、細胞の増殖・浸潤・転移を強めます[16]。ここでも、アクチンのヒスチジンメチル化(SETD3の産物)が足場として関わっている点は示唆に富みます。
6. CARNMT1──RNA代謝を根っこから支える修飾酵素
CARNMT1は、もともと筋肉に多いジペプチド「カルノシン」のヒスチジンをメチル化して「アンセリン」を作る酵素として知られていました。ところが近年の網羅的な解析で、CARNMT1が単なる小分子の修飾酵素にとどまらず、哺乳類における主要なタンパク質N1(π)位ヒスチジンメチル基転移酵素であることがわかりました[18]。CARNMT1が狙う目印は「Cx(F/Y)xH」という配列で、これはC3HジンクフィンガーというRNA結合タンパク質に特徴的な構造の中核をなします[17]。
💡 用語解説:ジンクフィンガー(亜鉛フィンガー)
亜鉛イオンを中心に、指のように突き出した小さな立体構造を作るタンパク質のパーツです。DNAやRNAをつかむ「手」として働きます。C3H型はシステイン3個とヒスチジン1個で亜鉛を挟むタイプで、多くのRNA結合タンパク質に見られます。CARNMT1はこの「ヒスチジンの部分」をメチル化することで、亜鉛のつかみ方やRNAとのくっつき方を微調整していると考えられます。
CARNMT1の相手には、RNA代謝のさまざまな段階を担う重要な因子が含まれます。スプライシング因子のU2AF1、mRNAの分解を制御するTTP(トリステトラプロリン)やRoquinなどです[17]。U2AF1は前駆体mRNAの3’スプライス部位を認識してスプライシングの組み立てを始める要の因子で、そのヒスチジンがCARNMT1にメチル化されると、RNAとの結合が最適化され、正しいスプライス部位の認識が保たれます[17]。CARNMT1を失った細胞では、U2AF1のRNA結合が広く乱れ、多数の選択的スプライシングに異常が生じます。しかもこの乱れは、酵素活性を失った変異型CARNMT1を入れても回復しないため、完全にヒスチジンメチル化という酵素活性に依存していることがわかっています[17]。
同じように、mRNAの安定性を制御するTTPやRoquinもCARNMT1の標的です。これらはmRNAの3’非翻訳領域(3’UTR)にある配列に結合して分解を促しますが、CARNMT1の活性が失われるとこれらの因子のRNA結合が妨げられ、炎症性サイトカインなど厳密な制御が必要な遺伝子の分解のタイミングが狂います[17]。こうしたRNA代謝の広範な制御は生命維持の根幹に関わり、CARNMT1を完全に欠いたマウス、あるいは酵素活性のない変異型だけを持つマウスは、いずれも胚性致死(胎児の段階で生存できない)という結果になりました[17]。
7. 「消す酵素」はあるのか──脱メチル化をめぐる議論
🔍 関連記事:リジン脱メチル化酵素/KDM6A遺伝子/エピジェネティクス治療
修飾が環境に応じて切り替わる「スイッチ」として働くには、付ける酵素(ライター)だけでなく、外す酵素(消しゴム=脱メチル化酵素)も必要です。ヒストンのリジンやアルギニンのメチル化では、KDMやLSDといった脱メチル化酵素が多数見つかっており、エピジェネティクス治療の標的として臨床応用が進んでいます。ところが、ヒスチジンメチル化専用の脱メチル化酵素が体の中に確かに存在するのかは、いまだ活発に議論されているテーマです[4]。
候補の一つが、DNAやRNAのメチル基を外す働きで知られるAlkBファミリー(ALKBH群やFTO)です。これらは鉄とα-ケトグルタル酸に依存する酵素で、がんの進行にも関わりますが、一部のホモログはヒスチジンからメチル基を外す可能性が推測されているものの、腫瘍の代謝産物である2-ヒドロキシグルタル酸によって阻害されるなど複雑な制御下にあり、ヒスチジン専用の消しゴムとしての機能はまだ立証されていません[19]。もう一つの候補であるJmjCドメインを持つ酵素群(JMJD5/RIOX1など)は、結晶構造の解析から、酵素自身のヒスチジンが活性中心をふさいでいたり、必要なループ構造が欠けていたりして、本当に脱メチル化酵素として働くのか疑問が投げかけられています[20]。
現時点での見方としては、アクチンのHis73メチル化のように細胞骨格の物理的な性質を決める修飾は、外れにくく長持ちする「静的な修飾」である可能性が高いと考えられています。一方で、RNAスプライシング因子やシグナル因子の修飾は環境に応じて動的に制御されるべきで、未知の脱メチル化酵素が隠れている可能性は依然として高く、大規模な探索が続けられています[4]。
8. どうやって測るのか──検出を阻んだ「技術の壁」と突破口
ヒスチジンメチル化の研究が、同じくらい重要なリジンやアルギニンのメチル化より数十年も遅れた最大の理由は、大規模に検出する手段が長い間なかったことにあります[1]。リジンメチル化では、修飾を特異的につかまえる抗体を使って濃縮する方法が標準ですが、ヒスチジンメチル化を効率よく濃縮できる抗体は今も開発されておらず、細胞からの直接的な質量分析による網羅解析は難しいとされてきました[1]。
この壁を破ったのが、すでに蓄積されていた超高深度のヒトプロテオームデータを情報科学的に「掘り直す」戦略です。膨大な質量分析スペクトルの中から、ヒスチジンにメチル基1つ分(+14.016 Da)の精密な質量のずれを持つペプチドをアルゴリズムで探し出しました[1]。さらに、質量のずれだけでは間違いを排除できないため、メチル化ヒスチジンに特有のイモニウムイオンという断片の検出が、修飾部位をアミノ酸単位で確定する決め手になりました[1]。この手法でHeLa細胞を解析した結果、299か所ものヒスチジンメチル化部位が同定され、ヒスチジンメチル化がアクチンやミオシンなど一部の例外ではなく、細胞や組織に広く行き渡った普遍的な修飾であることが明確になりました[1]。しかもこれらの部位は、機能的に重要なリン酸化部位や既知の疾患関連変異と統計的に重なりやすく、ヒスチジンメチル化が細胞内シグナルの「制御ハブ」として働いている可能性を強く支持しています[1]。
9. 遺伝医療との接点──いまは「研究段階」だからこそ知っておきたいこと
🔍 関連記事:臨床遺伝専門医とは/エピジェネティクス入門/タンパク質メチル化
ここまで見てきたように、ヒスチジンメチル化は細胞骨格・翻訳・亜鉛のバランス・RNA代謝という根幹に関わり、がん・原発性難産・ウイルス感染・胚発生といった幅広い病態につながります。ただし、現時点でSETD3・METTL9・METTL18・CARNMT1が標準的な遺伝子検査や保因者スクリーニングの対象になっているわけではありません。これらはまだ主に基礎研究・創薬研究の段階にあるテーマです。遺伝診療の現場では、こうした知見を「いま世界で何がわかりつつあるか」という文脈で理解し、確立した診断・治療とは切り分けて位置づけることが大切です。
遺伝子診断・遺伝形式・遺伝カウンセリングとの関わりで言えば、この分野の意義は次のように整理できます。第一に、SETD3とアクチンの研究は、原因のはっきりしない分娩の異常(原発性難産)を分子レベルで理解する手がかりになり得ます[8]。第二に、METTL9やMETTL18は将来の抗がん・抗転移薬の標的として、精密医療(プレシジョン・メディシン)の発展に関わります。そして第三に、これらはいずれもエピジェネティクス/翻訳後修飾という、遺伝情報の「使われ方」を制御する層の理解を深めるものです。タンパク質メチル化全体の中で、ヒスチジンメチル化がどこに位置づくのかを知っておくと、こうした新しい知見を受け止めやすくなります。
💡 豆知識:健康診断でおなじみの「3-メチルヒスチジン」
じつはヒスチジンメチル化は、身近な検査ともつながっています。アクチンやミオシンが分解されると、そこに含まれていた3-メチルヒスチジンが遊離しますが、これは再利用されずにそのまま排泄されます。このため尿中・血中の3-メチルヒスチジンは、筋肉のタンパク質がどれだけ分解されているか(筋タンパク質の代謝回転)を映す指標として研究・栄養評価の分野で使われています。ヒスチジンメチル化という一見マニアックな修飾が、「筋肉の状態を知る物差し」として役立っているわけです。
10. よくある誤解
誤解①「メチル化=DNAメチル化のことでしょう?」
よく知られたDNAメチル化は「DNAの塩基」に付く修飾です。一方この記事の主役は「タンパク質のアミノ酸」に付くメチル化で、まったく別の現象です。同じ『メチル化』でも、DNA・RNA・タンパク質のどこに付くかで意味が大きく異なります。
誤解②「珍しいマニアックな修飾だ」
かつてはアクチンなど一部のタンパク質だけの現象と思われていました。しかし網羅的な解析で数百か所の修飾部位が見つかり、全メチル化の約13%を占める、決して珍しくない修飾であることがわかっています。
誤解③「病気の遺伝子検査ですぐ調べられる」
現時点では、ヒスチジンメチル化を担う酵素は標準的な遺伝子検査や保因者検査の項目ではありません。あくまで基礎研究・創薬研究の段階にあるテーマで、臨床応用にはさらなる研究が必要です。
誤解④「メチル基が付くのはいつも良いこと」
修飾の意味は文脈しだいです。翻訳の品質を守る(METTL18)ように守りに働くこともあれば、がん細胞を細胞死に強くする(METTL9)ように病気側に働くこともあります。「良い・悪い」で単純に語れない仕組みです。
よくある質問(FAQ)
🏥 遺伝性疾患・遺伝子診断のご相談
遺伝性疾患や遺伝子検査、遺伝カウンセリングに関するご相談は
臨床遺伝専門医が在籍するミネルバクリニックに
お気軽にお問い合わせください。
参考文献
- [1] Large-scale identification of protein histidine methylation in human cells. NAR Genomics and Bioinformatics. [PMC8140740]
- [2] Protein Histidine Methylation. PubMed. [PubMed 32188384]
- [3] Molecular basis for METTL9-mediated N1-histidine methylation. PMC. [PMC10073072]
- [4] Enzymology and significance of protein histidine methylation. Journal of Biological Chemistry (PMC). [PMC8446795]
- [5] The Structure, Activity, and Function of the SETD3 Protein Histidine Methyltransferase. Life (Basel). [PMC8537074]
- [6] An engineered variant of SETD3 methyltransferase alters target specificity from histidine to lysine methylation. PMC. [PMC7049955]
- [7] Structural insights into SETD3-mediated histidine methylation on β-actin. eLife. [eLife 43676]
- [8] SETD3 is an actin histidine methyltransferase that prevents primary dystocia. PubMed. [PubMed 30626964]
- [9] Structure-function analysis of enterovirus protease 2A in complex with its essential host factor SETD3. PubMed. [PubMed 36075902]
- [10] siRNA screening identifies METTL9 as a histidine Nπ-methyltransferase that targets the proinflammatory protein S100A9. PMC. [PMC8571522]
- [11] METTL9-mediated N1-histidine methylation of zinc transporters promotes tumor growth. PMC. [PMC8674392]
- [12] METTL9-mediated N1-histidine methylation of SLC39A7 confers ferroptosis resistance and inhibits adipogenic differentiation in mesenchymal stem cells. PubMed. [PubMed 40414869]
- [13] Elevated METTL9 is associated with peritoneal dissemination in human scirrhous gastric cancers. PMC. [PMC8983939]
- [14] METTL18-mediated histidine methylation of RPL3 modulates translation elongation for proteostasis maintenance. eLife. [eLife 72780]
- [15] Human METTL18 is a histidine-specific methyltransferase that targets RPL3 and affects ribosome biogenesis and function. PMC. [PMC8034639]
- [16] METTL18 functions as a Phenotypic Regulator in Src-Dependent Oncogenic Responses of HER2-Negative Breast Cancer. PMC. [PMC11414398]
- [17] Histidine N1-position-specific methyltransferase CARNMT1 targets C3H zinc finger proteins and modulates RNA metabolism. PMC. [PMC10546975]
- [18] Putting a finger on histidine methylation. PMC. [PMC10546973]
- [19] The role of demethylase AlkB homologs in cancer. PMC. [PMC10060643]
- [20] Discovery and structural studies of histone demethylases. Frontiers in Epigenetics and Epigenomics. [Frontiers]






