目次

📚 入門シリーズ
この記事は「遺伝病を理解するためのヒトゲノム入門編」の1本です。全記事の一覧と学ぶ順番はヒトゲノム入門編・索引ページからご覧いただけます。
📍 クイックナビゲーション
遺伝子は、ただ持っているだけでは意味を持ちません。「いつ・どこで・どのくらい働かせるか」を決める調節のしくみがあって、はじめて体が正しく作られます。その調節の主役が「転写因子(てんしゃいんし)」です。この記事では、転写と翻訳の基本的な流れから、エンハンサーや3Dゲノムという最新の調節のしくみ、そして「遺伝子の設計図そのものは正常なのに病気が起こる」という不思議な遺伝病、さらに転写因子を狙う新しい薬まで、遺伝専門医が一般の方にもわかりやすく整理します。
Q. 転写因子とは何ですか?まず結論だけ知りたいです
A. 転写因子とは、DNAの特定の場所にくっついて、その近くの遺伝子を「オン」または「オフ」に切り替えるタンパク質のことです。いわば遺伝子の「スイッチ役」で、どの細胞でどの遺伝子を働かせるかを決めています。転写因子やその調節のしくみに異常が起こると、設計図(遺伝子の本体)が正常でも遺伝病が生じることがあり、近年はこの転写因子そのものを狙う新しい薬の開発も進んでいます。
- ➤転写の基本 → DNAの情報はまずRNAへ「転写」され、次にタンパク質へ「翻訳」される
- ➤調節のしくみ → エンハンサーや3Dゲノム(TAD・ループ)が遠くから遺伝子を操る
- ➤半分でも足りない → ハプロインサフィシエンシーという用量の問題が多くの遺伝病を説明する
- ➤配列は正常でも病気 → 遠くのエンハンサーやTAD境界の破綻で起こる「エンハンサー病・TAD病」
- ➤新しい治療 → 従来「狙えない」とされた転写因子を分解するPROTAC・分子接着剤
1. 転写因子とは?遺伝子の「オン・オフ」を決めるスイッチ役
私たちの体は、たった一つの受精卵から始まり、神経細胞・筋肉細胞・皮膚細胞など、姿も働きもまったく異なる約200種類の細胞へと分かれていきます。ところが、そのすべての細胞が持っている遺伝子(設計図)は基本的に同じです。それなのに細胞ごとに個性が生まれるのは、「どの遺伝子を働かせ、どの遺伝子を眠らせておくか」という組み合わせが細胞ごとに違うからです。この「働かせる・眠らせる」を実際に切り替えているのが転写因子です。
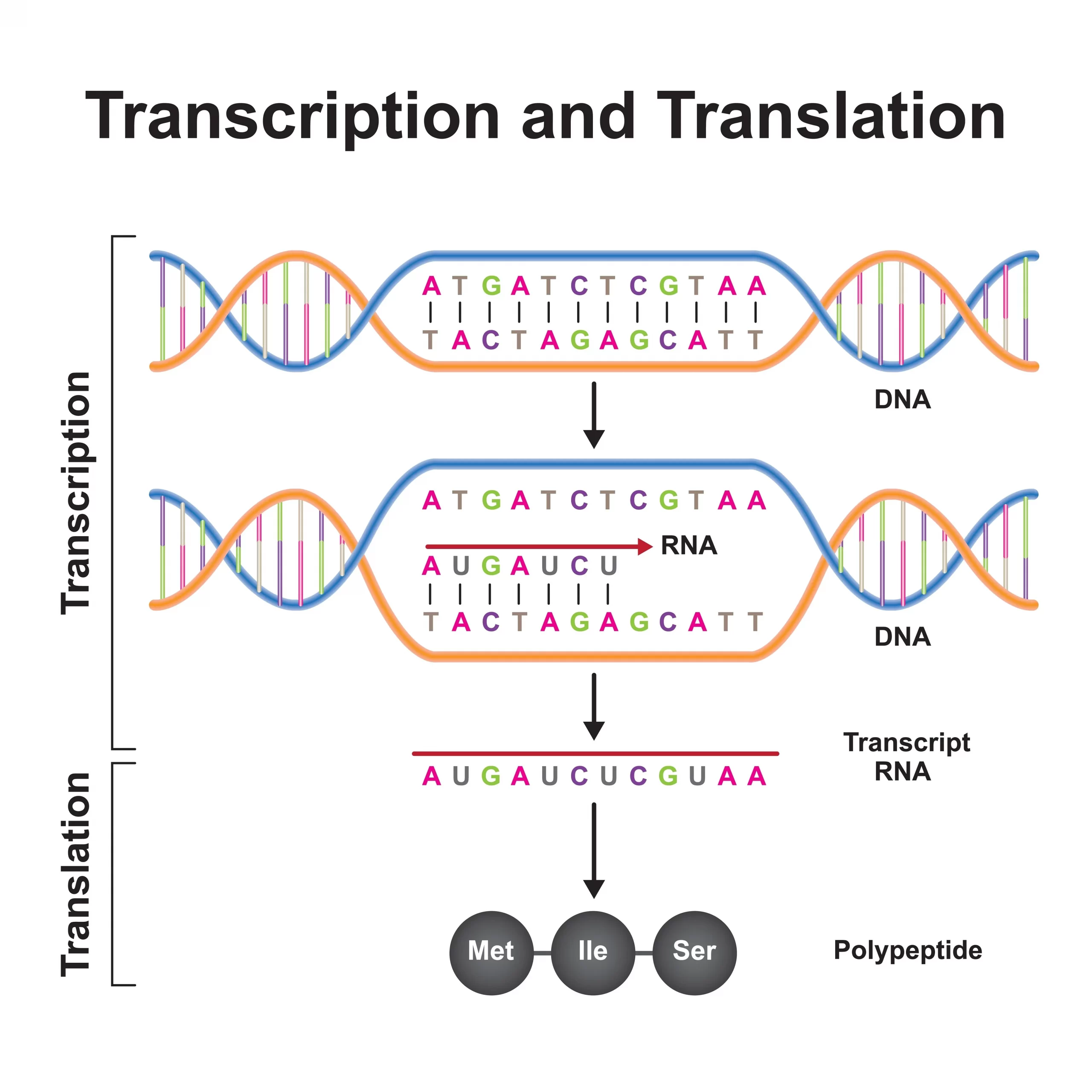
遺伝子が実際にはたらくまでの流れは、大きく二段階に分かれます。まずDNAの情報がRNA(メッセンジャーRNA)に写し取られる「転写(てんしゃ)」、次にそのRNAをもとにタンパク質が組み立てられる「翻訳(ほんやく)」です。この一方向の情報の流れは「セントラルドグマ」と呼ばれます。転写のより詳しい入門は遺伝子の発現 その1:転写概論、全体像はDNAからタンパク質へで解説しています。
セントラルドグマと転写の向き
RNA合成
タンパク合成
転写のとき、RNAポリメラーゼは鋳型(型紙)となるDNA鎖を3’→5’の向きに読み取り、新しいRNAは5’→3’の向きに伸ばしていきます。この「向き(方向性)」は決まっており、逆流はしません。
この転写を始めるには、RNAポリメラーゼという酵素が、遺伝子の入り口にあたるプロモーターという領域に取りつく必要があります。ヒトなど真核生物では、ポリメラーゼが単独で取りつくのではなく、複数のタンパク質が集まって足場を作り、そこにポリメラーゼが乗り込みます。この足場作りや、遺伝子を強めたり弱めたりする調節を担うのが転写因子です。多くの転写因子は、DNAの特定の「文字の並び(配列)」を読み取るDNA結合ドメインという部品を持ち、そこで狙った場所にピンポイントで結合します。
💡 用語解説:転写因子とプロモーター
転写因子は、DNAに結合して近くの遺伝子の転写量を上げたり下げたりするタンパク質です。ポリメラーゼを呼び込む「基本転写因子」と、特定の遺伝子だけを組織や状況に応じて調節する「特異的転写因子」に大きく分けられます。
プロモーターは遺伝子の「スタートライン」にあたる領域です。真核生物では「TATAボックス」と呼ばれる目印の配列や、CとGが多いCpGアイランドがしばしば存在し、ここのメチル化(後述)で遺伝子の働きが抑えられることもあります。詳しくはプロモーターの用語ページもご覧ください。
転写因子は「遺伝子を働かせるための最初の一手」を担うため、その働きが少しでも狂うと、体づくりや代謝、環境への適応など、あらゆる場面に影響が及びます。だからこそ、転写因子の異常は数多くの遺伝病やがんの背景にあり、そのしくみを理解することは、遺伝子診断や新しい治療を考えるうえで欠かせません。転写因子そのものの分類や構造については転写因子(総論)で詳しくまとめています。
2. エンハンサー:遠くから遺伝子を操る「調節スイッチ」
🔍 関連ページ:エンハンサー/スーパーエンハンサー/エンハンサーRNA(eRNA)
遺伝子の調節は、プロモーターの近くだけで完結するわけではありません。ヒトのゲノムには、タンパク質を作る遺伝子が約2万個しかないのに対し、遺伝子の働きを調節するエンハンサーの候補は100万か所以上もあると考えられています。エンハンサーは遺伝子から遠く離れた場所にあることも多く、そこに組織ごとに決まった転写因子が結合すると、離れた遺伝子の転写を大きく高めます。逆に働きを抑える領域は「サイレンサー」と呼ばれます。
エンハンサーは、いつでも同じ強さで働いているわけではありません。周囲のヒストン(DNAを巻き取るタンパク質)の目印や、結合している転写因子の状況によって、大きく三つの状態を行き来します。準備状態(Poised)は、いつでも動けるよう先回りの目印だけがついてスタンバイしている段階、活性化状態(Active)は実際に強く働いている段階、そして潜在状態(Latent)は、特定の刺激が来て初めて目覚める段階です。細胞は環境や発生段階に応じてこの状態を切り替えることで、必要な遺伝子だけを的確に働かせています。
かつては「意味のない副産物」と思われていたエンハンサーRNA(eRNA)という短いRNAも、近年ではエンハンサーとプロモーターを物理的に近づける橋渡しを安定させ、活性化状態を保つのに一役買っていることが分かってきました。また、活発なプロモーターの約4分の1は、別の遠くの遺伝子に対してはエンハンサーとしても働くという「二役」を担っており、ゲノムの調節は想像以上に立体的で柔軟です。とりわけ多数のエンハンサーが密集したスーパーエンハンサーは、細胞の運命を決める重要遺伝子を強力に制御します。
3. 3Dゲノムとループ押し出し:CTCFとコヒーシンが作る立体構造
🔍 関連ページ:TAD(トポロジカル関連ドメイン)/ループ押し出し/コヒーシン
エンハンサーが遠くの遺伝子を操れるのは、DNAが細胞の核の中で折りたたまれ、遠い場所どうしが近づける立体構造を作っているからです。この折りたたみの基本単位がTAD(トポロジカル関連ドメイン)です。TADは「ご近所づきあいの範囲」を仕切る区画のようなもので、同じTADの中にあるエンハンサーと遺伝子は活発にやり取りしますが、TADの壁(境界)を越えた相手とは基本的に交流しません。染色体全体の折りたたみについては染色体の構造もあわせてご覧ください。
このTADやループがどう作られるのかを説明するのがループ押し出しという考え方です。リング状のタンパク質コヒーシンが、NIPBLという補助因子によってDNAに載せられ、DNAを両側からたぐり寄せながらループをどんどん大きくしていきます[5]。そして、DNA結合タンパク質CTCFが向かい合わせ(収束配向)に結合した場所に来ると、コヒーシンの前進が止まり、そこがループの「留め具」=TADの境界になります[6]。CTCFはコヒーシンにブレーキをかけるだけでなく、進む向きをそろえる役割も担っていることが、一分子レベルの観察から明らかになっています[6]。
ループ押し出しでTADができるまで
① 積み込み
NIPBLがコヒーシン(リング)をDNAに載せる
② たぐり寄せ
コヒーシンがDNAを両側から通し、ループを拡大
③ 停止=境界
向かい合ったCTCFで停止し、ループの留め具(TAD境界)に
💡 用語解説:TAD・コヒーシン・CTCF
TADは、DNAの折りたたみで作られる「ご近所区画」です。コヒーシンはDNAをたぐり寄せてループを作るリング状のタンパク質、CTCFはそのループの端に立つ「杭(くい)」のような目印です。
この立体構造が正しく保たれることで、エンハンサーは「担当の遺伝子」だけを適切に調節できます。区画の壁が壊れると、本来関係のないエンハンサーが別の遺伝子に手を出してしまい、病気につながります(後述のTAD病)。コヒーシンの要となるRAD21や積み込み役のNIPBLの異常は、コルネリア・デ・ランゲ症候群などの原因となります。
さらに近年注目されているのが、液–液相分離(LLPS)というしくみです。水の中に油滴が浮かぶように、転写に必要な因子が核内の一か所に「液滴」のように集まり、遺伝子を集中的に読み出す作業場を作ります。CTCFが作る立体的なループは、この作業場を正しい場所に配置するための土台になっていると考えられており、立体構造・相分離・転写因子が三位一体で遺伝子発現を支えていることが分かってきました。
4. ハプロインサフィシエンシー:遺伝子が「半分」でも病気になる理由
🔍 関連ページ:ハプロ不全(ハプロインサフィシエンシー)/ハプロ不全予測指標(%HI)
ヒトは同じ遺伝子を父由来・母由来の2本持っています。転写因子やクロマチンを調節する遺伝子では、片方が壊れて働く量が半分(50%)になっただけで病気になることが少なくありません。これをハプロインサフィシエンシー(ハプロ不全)=「半量では足りない」現象と呼びます。多くが片方の変異だけで症状が出るため、遺伝の形式としては常染色体顕性(優性)遺伝の形をとります。
なぜ半分では足りないのでしょうか。細胞が「神経になる/筋肉になる」といった運命を決めるとき、転写因子は自分自身の量をさらに増やす正のフィードバックや、複数がチームを組んで一気にスイッチを入れる協調的な結合を利用しています。これによって、遺伝子は中途半端な状態ではなく「オン」か「オフ」かをはっきり決める、スイッチのような応答を示します。ところが転写因子の量が半分に減ると、このスイッチを押し切る「しきい値」に届かなくなり、システムが「オン」を維持できずに崩れてしまうのです。これが、同じ変異でも症状の重さに大きな個人差が出たり、症状がはっきり出ない人(浸透度が不完全)がいたりする理由の一つと考えられています。用量の感受性は%HIという予測指標で見積もることもできます。
5. エンハンサー病・TAD病:設計図は正常でも起こる遺伝病
遺伝病というと「遺伝子そのものに傷がある」とイメージされがちですが、実は遺伝子の本体はまったく正常なのに、その調節領域や立体構造の壊れ方だけで病気になることがあります。ゲノム全体で見ると、病気と関わる遺伝的な個人差の多くは、タンパク質を作らない「非コード領域」にあることが分かっており、調節のしくみの理解が診断の鍵になります。
その代表例が、目の虹彩ができない無虹彩症です。原因遺伝子であるPAX6の本体が無傷でも、そこから約150キロ塩基も離れた「SIMO」という超保存エンハンサーに小さな点変異が起こるだけで、目の発生に必要なPAX6の発現が維持できなくなり、無虹彩症が生じます[1][2]。これは「遺伝子の設計図」ではなく「調節スイッチ」の故障で起こる病気であり、こうしたタイプはエンハンサー病と呼ばれます。
似た例に、手足の形態を決めるSHHがあります。SHHの手足での発現を制御するエンハンサー「ZRS」は、SHH遺伝子から約100万塩基(1メガベース)も離れた場所にあります。このZRSに変異が入ると、本来は出ないはずの部位でSHHが誤って発現し、指が余分にできる前軸多指症などが生じます[3]。SHH遺伝子そのものは正常でも、遠くのスイッチの誤作動だけで手足の形が変わってしまうのです。
一方、TADの「壁」が壊れて起こるのがTAD病です。区画を仕切る境界が欠失・逆位・重複でこわれると、本来は別区画にいたエンハンサーが隣の遺伝子に手を出し、間違った場所・時期に遺伝子が働いてしまいます。実際に、指や手足の奇形の一部は、遺伝子ではなくTAD境界の構造変化によって、エンハンサーと遺伝子の「配線」が組み替わることで起こると証明されています[4]。この「エンハンサーの乗っ取り」はエンハンサー・ハイジャッキングとも呼ばれ、一部のがんの発症にも関わります。
このほか、下垂体機能低下や性発達に関わるSOX3、耳の形成に関わるHOXA2(無耳症・小耳症)、言語に関わるFOXP2、頭蓋骨が早く癒合するTFAP2Bなど、多くの転写因子でハプロ不全や調節異常が知られています。共通するのは、「遺伝子の量」や「調節の配線」がわずかに狂うだけで、体づくりの特定の部分に集中して影響が出るという点です。こうした遠隔調節やゲノムの区画の破綻は、まとめてゲノム病という枠組みでも整理されます。
6. 多因子疾患・加齢・エピジェネティクスと転写
🔍 関連ページ:GWAS(ゲノムワイド関連解析)/DNAメチル化/エピジェネティクス
転写の調節異常は、一つの遺伝子で決まる病気だけでなく、環境と体質が絡み合う多因子疾患や、年齢を重ねることで起こる病気とも深く関わります。たとえば自己免疫疾患である多発性硬化症(MS)では、病気のなりやすさに関わる遺伝的な個人差の多くが、免疫細胞の炎症スイッチであるNF-κB経路の調節領域に集まっていることが分かっています[9]。こうした変異を持つ人では、まだ活性化していないナイーブなCD4陽性T細胞の段階からNF-κBが過剰に働きやすく、感染などの環境刺激をきっかけに過剰な免疫反応が引き起こされると考えられています[10]。これは、遺伝子の個人差がGWASという手法で「炎症の調節スイッチ」として浮かび上がってきた好例です。
先天性心疾患や口唇口蓋裂でも、心臓や顔の発生を指揮するマスター転写因子(NKX2.5・GATA4・TBX5など)のDNA結合ドメインに新生突然変異(de novo変異)が起こり、下流の遺伝子プログラムがうまく起動しなくなることが主な原因の一つとされています。こうした変異は、妊娠中の母体の環境要因(糖尿病や特定の薬剤・化学物質への曝露など)と相互に作用して、発生の破綻を後押しすることがあります。
💡 用語解説:ミスセンス変異とde novo変異
ミスセンス変異とは、遺伝子の1文字が変わることで、作られるタンパク質のアミノ酸が1つ別のものに置き換わる変化です。転写因子のDNA結合ドメインにこれが起こると、DNAをうまくつかめなくなり、遺伝子の調節に支障が出ます。
de novo変異(新生突然変異)は、両親のどちらにもなく、その子で初めて生じた変化です。家族歴がなくても発症しうるのはこのためで、遺伝カウンセリングでは「なぜうちの家系に」という疑問に答える重要な概念になります。
加齢との関わりでは、幹細胞の自己複製を支えるSOXファミリー転写因子の働きが、酸化ストレスや慢性炎症、DNAメチル化などのエピジェネティックな乱れによって低下することが知られています。DNAメチル化は、DNAの文字を変えずに遺伝子の働きを抑える「付箋」のようなしくみで、ヒストン修飾とともに、多くの転写因子の結合しやすさを左右します。また、増殖の司令塔である転写因子MYCは、いくつもの化学修飾のリレーによって寿命が精密に管理されており、その分解の合図が失われると、下流の遺伝子が際限なく暴走してしまいます。エピジェネティクスの全体像はエピジェネティクスの回で詳しく扱います。
7. 転写因子を狙う創薬:PROTACと分子接着剤
🔍 関連ページ:標的タンパク質分解(TPD)/PROTAC/分子接着剤
転写因子は多くの病気の中心にありながら、長らく「薬で狙えない標的(アンドラッガブル)」の代表とされてきました。従来の薬は、酵素などの「くぼみ(ポケット)」にはまり込んで働きを止めますが、転写因子はぐにゃぐにゃと形が定まらない部分が多く、はまり込む場所が乏しいためです。この壁を突破したのが、細胞にもともと備わる「不要なタンパク質を分解するしくみ(ユビキチン・プロテアソーム系)」を逆に利用して、標的そのものを丸ごと壊してしまう標的タンパク質分解(TPD)という発想です。
💡 用語解説:PROTACと分子接着剤(molecular glue)
PROTAC(プロタック)は、「標的にくっつく部分」と「分解装置(E3リガーゼ)を呼ぶ部分」を、ひもでつないだ二股のキメラ分子です。標的と分解装置を物理的に橋渡しして、標的にゴミ処理の目印(ユビキチン)を付けさせ、分解へと導きます。
分子接着剤(molecular glue)は、より小さな一つの分子で、分解装置の表面をわずかに変形させ、本来は無関係だった標的を「のり付け」して分解を誘導します。小さいぶん体内に入りやすいのが特長です。
この二つは、性格の異なる技術として発展してきました。多発性骨髄腫の治療薬レナリドミド(サリドマイドの誘導体)は、代表的な分子接着剤です。分解装置CRBNにくっつくことで、骨髄腫の生存に必須の転写因子IKZF1・IKZF3を選び出して分解させる——という、当初は謎だった作用が解明されました[7]。一方PROTACでは、去勢抵抗性前立腺がんのアンドロゲン受容体を狙うARV-110(バブデガルタミド)や、乳がんのエストロゲン受容体を狙うARV-471が、いずれも飲み薬として臨床試験で評価されてきました[8]。従来は狙えなかった転写因子を「壊す」という戦略が、実際の患者さんへ届き始めています。
さらに近年は、構造解析や計算科学の進歩により、偶然に頼らず「狙って作る」次世代の分子接着剤の研究も進んでいます。たとえば、特定の化学構造を目印に用いた新しい分子接着剤に、ナノ粒子の技術を組み合わせ、正常な組織を傷つけずに腫瘍のある場所でだけ薬を放出させる——といったアプローチが研究段階で報告されています。転写調節を標的とする治療は、これからの精密医療の重要な柱の一つになると期待されています。

8. 転写因子の異常と遺伝診療:どこで臨床とつながるのか
🔍 関連ページ:遺伝カウンセリングとは/臨床遺伝専門医とは
転写因子とその調節は、基礎研究の話にとどまりません。実際の遺伝診療では、この理解が「診断」と「遺伝カウンセリング」の両面で役立ちます。診断面では、症状に合う遺伝子の本体を調べても変化が見つからないとき、エンハンサーやTAD境界といった非コード領域まで視野を広げる必要があると分かります。頭蓋骨が早く癒合する病気(頭蓋骨縫合早期癒合症)のように、複数の転写因子や経路が関わる疾患では、関連遺伝子をまとめて調べるNGSパネル検査が診断の助けになります。
遺伝カウンセリングの面では、ハプロ不全という考え方が特に重要です。片方の遺伝子の変化だけで発症しうること、しかし浸透度や重症度に幅があること、多くがde novo変異で家族歴なく起こりうること——これらを正確に伝えることで、ご家族は「次の妊娠でどうなるのか」「本人の子にどう受け継がれるのか」を現実的に理解できます。当院ではこうした説明を、臨床遺伝専門医が担当します。転写因子は「未来をすべて決める設計図」ではなく「幅のある調節スイッチ」であるという理解は、結果の受け止め方そのものを支えてくれます。
💡 押さえておきたいポイント
遺伝子の「設計図(本体)」が正常でも、調節スイッチ(エンハンサー)や立体構造(TAD)の異常だけで遺伝病が起こりえます。検査で「本体に異常なし」と言われても、調節領域の可能性が残る場合があるため、症状と合わない結果のときは遺伝カウンセリングで解釈を整理することが大切です。
9. よくある誤解
誤解①「遺伝子に異常がなければ遺伝病ではない」
遺伝子の本体が正常でも、遠くのエンハンサーやTAD境界という調節のしくみの異常だけで遺伝病が起こることがあります。無虹彩症や一部の手足の奇形がその例です。
誤解②「片方の遺伝子が正常なら大丈夫」
転写因子では、片方が正常でも残りの半分の量では足りない(ハプロ不全)ことがあります。多くが片方の変異だけで発症し、常染色体顕性(優性)遺伝の形をとります。
誤解③「同じ変異なら症状も同じ」
転写因子はスイッチのように応答するため、しきい値ギリギリの人ではわずかな差で結果が大きく振れます。同じ変異でも重症度や浸透度に幅が出るのはこのためです。
誤解④「転写因子は薬で狙えない」
かつてはそう考えられていましたが、PROTACや分子接着剤という「壊す」戦略により、転写因子を標的とする薬の研究・臨床開発が現実に進んでいます。
よくある質問(FAQ)
🧬 遺伝子・遺伝についてのご相談
遺伝子検査の結果の見方や、遺伝の形式・再発リスクなど
遺伝に関するご不安は、臨床遺伝専門医が在籍する
ミネルバクリニックにお気軽にご相談ください。
参考文献
- [1] Bhatia S, et al. Disruption of autoregulatory feedback by a mutation in a remote, ultraconserved PAX6 enhancer causes aniridia. Am J Hum Genet. 2013. [PubMed 24290376]
- [2] OMIM #617141. Aniridia 2 (AN2). Johns Hopkins University. [OMIM 617141]
- [3] Lettice LA, et al. A long-range Shh enhancer regulates expression in the developing limb and fin and is associated with preaxial polydactyly. Hum Mol Genet. 2003. [PubMed 12837695]
- [4] Lupiáñez DG, et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell. 2015. [PubMed 25959774]
- [5] Kim Y, et al. Human cohesin compacts DNA by loop extrusion. Science. 2019. [Science]
- [6] Davidson IF, et al. CTCF is a DNA-tension-dependent barrier to cohesin-mediated loop extrusion. Nature. 2023. [Nature]
- [7] Krönke J, et al. Lenalidomide causes selective degradation of IKZF1 and IKZF3 in multiple myeloma cells. Science. 2014. [Science]
- [8] Arvinas, Inc. PROTAC protein degrader bavdegalutamide (ARV-110) and ARV-471 clinical data. [Arvinas]
- [9] International Multiple Sclerosis Genetics Consortium. Low-Frequency and Rare-Coding Variation Contributes to Multiple Sclerosis Risk. Cell. 2018. [Cell]
- [10] Nuclear Factor κB (NF-κB)–Mediated Inflammation in Multiple Sclerosis. Frontiers in Immunology. 2020. [Frontiers]






