目次

ARL13B(ADP-ribosylation factor-like 13B)は、全身のほぼあらゆる細胞に存在するアンテナ状の感覚小器官「一次繊毛」の内部に特異的かつ高濃度に局在する低分子量GTPアーゼです。脳内では分子マーカーとして広く活用されるだけでなく、近縁タンパク質ARL3の活性化因子(GEF)として脂質修飾タンパク質の精密な繊毛内輸送を担い、Sonic Hedgehog(Shh)シグナル伝達を繊毛の「外部」から遠隔制御するという驚くべき機能まで持ちます。変異によってジュベール症候群(繊毛病)を引き起こし、過剰発現はグリオーマや胃がんの悪性化に関与することも明らかになっています。
Q. ARL13B遺伝子とはどのような遺伝子ですか?まず結論だけ知りたいです
A. 一次繊毛の微小管軸糸と繊毛膜に特異的かつ高濃度に局在する低分子量GTPアーゼをコードする遺伝子です。近縁タンパク質ARL3のGEF(活性化因子)として脂質修飾タンパク質の繊毛内精密輸送を司り、Shhシグナル伝達・大脳皮質発生・網膜形成・精子形成など多彩な生理的役割を担います。変異によってジュベール症候群(繊毛病)を引き起こします。
- ➤遺伝子の概要 → Rasスーパーファミリー・ARFファミリーに属する進化的に高度に保存されたGTPアーゼ遺伝子
- ➤分子構造 → N末端パルミトイル化・VxPxモチーフによる繊毛特異的局在、非定型的C末端ドメイン
- ➤GEFカスケード → ARL3を活性化して脂質修飾カーゴ(INPP5Eなど)を繊毛内へ精密ターゲティング
- ➤Shhシグナルのパラダイムシフト → V358A変異モデルが「繊毛外からの遠隔制御」を証明
- ➤関連疾患 → ジュベール症候群(繊毛病)・グリオーマ・胃がん・増殖性硝子体網膜症
1. ARL13B遺伝子・タンパク質の概要:進化的背景と生物学的意義
ARL13B(ADP-ribosylation factor-like 13B)は、Rasスーパーファミリー内のARF(ADP-ribosylation factor)ファミリーに属する低分子量GTPアーゼの一種で、特異かつ非定型的な特徴を持つ重要なメンバーです。この遺伝子は進化的に極めて高度に保存されており、真核生物の最後の共通祖先にすでに存在していたとされています。進化の過程において、尾索動物の段階でARL13AとARL13Bへの遺伝子重複が生じ、その後それぞれ異なる機能へと特化してきました。
哺乳類をはじめとする多くの脊椎動物において、ARL13Bはほぼすべての細胞型に存在するアンテナ状の感覚小器官「一次繊毛」の微小管軸糸および繊毛膜に特異的かつ高濃度に局在します。脳内においては、アデニル酸シクラーゼ3(ADCY3)と並ぶ「一次繊毛の確立された分子マーカー」として広く活用されています。
しかしARL13Bは単なる構造マーカーにとどまりません。繊毛の構造的完全性の維持(シリオジェネシス)、微小管に基づく軸糸構造の制御、細胞内タンパク質輸送の空間的オーケストレーション、そして胚発生から成体の恒常性維持に至るまでの多彩なシグナル伝達において、極めて動的かつ中心的な役割を果たすマスターレギュレーターです。遺伝学的な観点からは、ARL13Bの変異は重篤な神経発達障害「ジュベール症候群(Joubert Syndrome: JS)」の直接的な原因となり、繊毛病(シリオパシー)の主要な病因遺伝子としての地位を確立しています。
💡 用語解説:一次繊毛(プライマリー・シリア)とは
一次繊毛とは、ほぼすべての哺乳類細胞の表面に1本だけ生えている、髪の毛のような超微細な突起状の細胞小器官です(長さ:約1〜10マイクロメートル)。かつては退化した器官とみなされていましたが、現在では細胞外の環境変化を感知する「アンテナ」として、多種多様なシグナル伝達経路の制御拠点として機能することが明らかになっています。内部は微小管でできた「軸糸(axoneme)」と呼ばれる骨格構造で支えられており、ARL13Bはこの軸糸および周囲の繊毛膜に特異的に濃縮して存在します。一次繊毛の機能不全は「繊毛病(シリオパシー)」と呼ばれる疾患群を引き起こします。
💡 用語解説:低分子量GTPアーゼとは
GTPアーゼとは、GTP(グアノシン三リン酸)をGDP(グアノシン二リン酸)に加水分解することができる酵素の総称です。低分子量GTPアーゼ(別名:Gタンパク質、スモールGプロテイン)は分子量約20〜30kDaの小型タンパク質で、GTP結合時=「オン」(活性型)・GDP結合時=「オフ」(不活性型)というスイッチのように働きます。細胞内のシグナル伝達・物質輸送・細胞骨格制御などに広く関与しています。Rasスーパーファミリーの代表例としては、がんとの関連で知られるRasタンパク質や、物質輸送を担うRabタンパク質などがあります。ARL13BはARF(ADP-ribosylation factor)ファミリーに分類されます。
2. 非定型的な分子構造と生化学的特性
ARL13Bは他のARFファミリーGTPアーゼと比較して、著しく異なる非定型的なドメイン構造と生化学的特性を有しています。この構造的特異性が、一次繊毛という特殊な細胞内コンパートメントにおける複雑な機能発現を可能にしています。
主要な構造的特徴と機能の一覧
🔴 N末端パルミトイル化
疎水性相互作用を介した繊毛膜・細胞膜への強固なアンカリングに必須。この脂質修飾によってARL13Bは繊毛膜と安定した会合を形成し、界面活性剤で処理しないと繊毛から可溶化されない。
🔵 Gドメイン(GTP結合領域)
GTP/GDPサイクルによる活性状態のスイッチング。微小管軸糸への物理的アンカリングと、ARL3に対するGEF活性の構造的基盤。Gドメイン欠損変異体は繊毛内保持率が有意に低下(p=0.028)。
🟣 非定型C末端ドメイン(約20kDa)
他のGTPアーゼには見られない特有の伸長ドメイン。繊毛内局在の長期的維持、エフェクタータンパク質との相互作用、および全体的な機能発現に不可欠。この領域の欠失は機能不全を招く。
🟢 VxPxモチーフ
繊毛へのターゲティングを規定する局在化シグナル配列。このモチーフを特異的に破壊するV358A点変異を導入すると、タンパク質は繊毛内から完全に排除される。Shhシグナル研究の鍵となるモチーフ。
🟡 RVEPモチーフ
繊毛特異的輸送アダプタータンパク質(Rab8やTNPO1など)との相互作用インターフェース。細胞質から繊毛への効率的な輸送に必要な繊毛ターゲティング配列の一部として機能する。
💡 用語解説:パルミトイル化(Palmitoylation)とは
パルミトイル化とは、タンパク質のシステイン(アミノ酸の一種)残基にパルミチン酸(炭素16個の飽和脂肪酸)が共有結合的に付加される翻訳後修飾の一種です。この脂質修飾によってタンパク質は強い疎水性(水をはじく性質)を獲得し、細胞膜や特殊な膜構造(繊毛膜など)に安定して結合できるようになります。ARL13BのN末端パルミトイル化は、同タンパク質が繊毛膜に特異的かつ安定して局在するための最重要メカニズムです。パルミトイル化は可逆的な修飾であり、繊毛内でのARL13Bの動的な動態を可能にしています。
GTPアーゼ活性の特異性と主要な分子間相互作用
ARL13BはGTPに結合する能力を持つものの、一般的なGTPアーゼとは異なり、その内在性のGTP加水分解活性は極めて低いという特徴があります。精製されたマウス組み換えArl13bタンパク質を用いた生化学的アッセイにより、GTPの加水分解はARL13B単独ではほとんど進行せず、特異的なGTPアーゼ活性化タンパク質(GAP)の存在下において初めて強力に促進されることが証明されています。また、ARL13Bはカゼインキナーゼ2(Casein kinase 2)の基質にもなることが発見されており、リン酸化による機能的微調整の可能性も指摘されています。
一次繊毛内において、ARL13Bは単独で機能するのではなく、多数のタンパク質と複雑な相互作用ネットワークを形成します。ARL13Bはチューブリンと直接結合し、微小管に基づく軸糸構造の安定化に寄与します。また、繊毛内輸送の中枢機構であるIFT(繊毛内輸送:Intraflagellar Transport)複合体のサブコンプレックスAとBの間の物理的会合の維持にも関与しており、エキソシスト複合体やUNC119などのタンパク質輸送因子とも相互作用することが確認されています。
💡 用語解説:IFT(繊毛内輸送/Intraflagellar Transport)とは
IFTとは、一次繊毛の内部で物質を運ぶ特殊な輸送システムです。繊毛内には細胞体側から繊毛先端へ向かう「順行性IFT(キネシンモーター使用)」と、先端から細胞体へ向かう「逆行性IFT(ダイニンモーター使用)」の2方向の輸送があります。IFT複合体A(逆行性)とB(順行性)という2種類のサブコンプレックスが存在し、ARL13Bはこれらの複合体間の物理的な会合を維持することで、繊毛内でのタンパク質の双方向輸送を調節しています。IFTの異常も繊毛病の原因となります。
3. ARL13B→ARL3 GEFカスケード:脂質修飾タンパク質の繊毛内輸送システム
ARL13Bが担う最も重要かつ分子レベルで詳細に解明された生化学的機能が、細胞内局在を同じくする別のARF様GTPアーゼ「ARL3」に対するグアニンヌクレオチド交換因子(GEF:Guanine nucleotide Exchange Factor)としての役割です。このARL13B→ARL3カスケードは、脂質修飾を受けた多様なシグナル伝達タンパク質を一次繊毛の内部へと運び込み、正しい空間座標で放出するための高度な制御システムの中核を担います。
💡 用語解説:GEF(グアニンヌクレオチド交換因子)とは
GEF(Guanine nucleotide Exchange Factor)とは、不活性なGDP結合型GTPアーゼに結合してGDPを強制的に解離させ、代わりにGTPを結合させることでGTPアーゼを活性化するタンパク質です。細胞内にはGTPが豊富に存在するため、GEFがGDPを引き剥がすことでGTPアーゼは自動的に活性型(GTP結合型)になります。ARL13BはARL3に対して非常に高い基質特異性を持ち、近縁のARL2やARL6(BBS3)は活性化しません。このGEF活性は、ARL13B自身がGTP結合型(活性型)にあるときにのみ最大限発揮されるという自己制御機構も備えています。
GEF活性の速度論的データ
このGEF反応の速度は、生化学的キネティクスアッセイ(蛍光スペクトロメーターを用いmantGDPの解離を測定)によって精密に定量化されています。クラミドモナス(Chlamydomonas reinhardtii)の実験系において、ARL3からのGDPの自発的な解離速度(kobs)はわずか1.4 × 10⁻⁴ s⁻¹に過ぎませんが、GTP結合型の野生型ARL13Bが存在する環境下では0.91 × 10⁻² s⁻¹へと約65倍も劇的に加速します。このGEF活性の構造的基盤は、X線結晶構造解析(PDB: 5DI3、分解能2.50 Å)によって分子レベルで解明されており、ARL13BのGドメインに加えてC末端の特定のαヘリックス(α-6ヘリックス、残基212〜228)が触媒作用に不可欠であることが明らかになっています。
脂質修飾カーゴ輸送の4段階プロセス
ARL13B→ARL3カスケードの生理的目的は、ファルネシル化・ゲラニルゲラニル化・ミリストイル化といった疎水性の脂質修飾を受けたカーゴタンパク質(代表例:ホスファターゼINPP5E、腎疾患関連タンパク質NPHP3、視覚伝達タンパク質GNAT-1など)を、水溶性の細胞質から一次繊毛の内部へと安全に輸送・放出することです。
① カーゴのマスキングと輸送
細胞質内で脂質修飾されたカーゴタンパク質は、キャリアタンパク質(プレニル化カーゴ → PDE6δ、ミリストイル化カーゴ → Unc119a/b)と結合します。キャリアは疎水性の脂質基を覆い隠し(マスキング)、水溶性環境での輸送を可能にしたまま繊毛内へ移行します。
② 空間特異的な活性化(コンパートメント形成)
ARL3自体は細胞質全体に広く分布しますが、そのGEFであるARL13Bは一次繊毛膜上に排他的に局在します。このため、ARL3は「繊毛の内部に入ったときのみ」GTP結合型(活性型)に変換され、繊毛内は高濃度な「ARL3・GTPコンパートメント」として確立されます。
③ アロステリックなカーゴ放出
活性型ARL3・GTPは、キャリアタンパク質(PDE6δやUnc119)に対して極めて高い親和性を持ちます。ARL3・GTPがキャリアに結合することでアロステリックな構造変化が起こり、カーゴ(INPP5Eなど)がキャリアから解離します。放出されたカーゴは自身の脂質修飾基で繊毛膜に直ちに定着します。
④ シグナルの終焉とリサイクル
繊毛基部の移行ゾーン(トランジションゾーン)付近に存在するRP2(ARL3に特異的なGAP)がARL3・GTPをARL3・GDPに加水分解します。これによりARL3とキャリアタンパク質が細胞質にリサイクルされ、次の輸送サイクルに備えます。
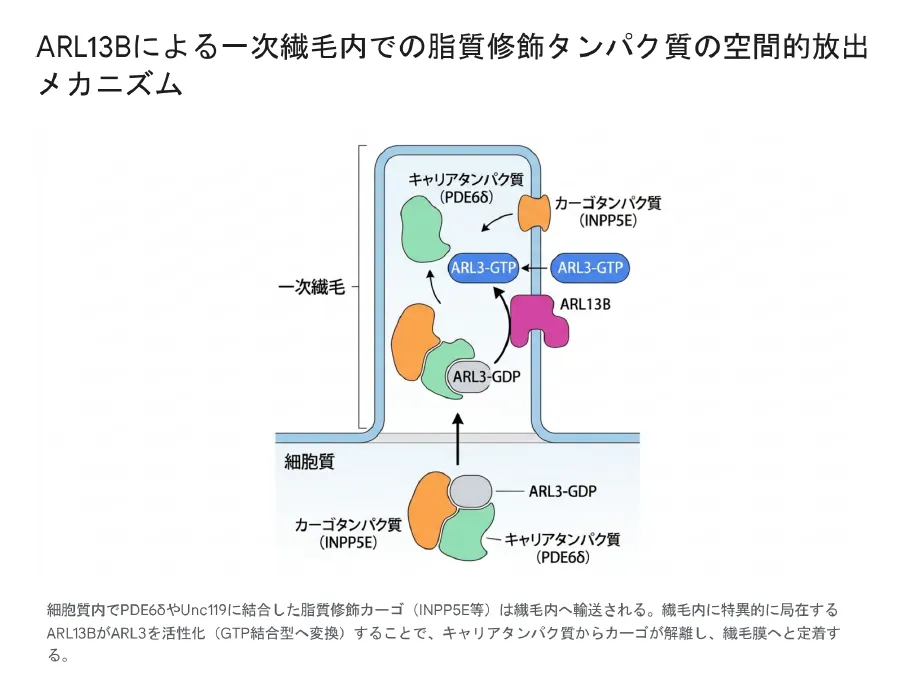
細胞質でPDE6δやUnc119に結合した脂質修飾カーゴ(INPP5E等)は繊毛内へ輸送される。繊毛内に特異的に局在するARL13BがARL3を活性化(GTP結合型へ変換)することで、キャリアタンパク質からカーゴが解離し繊毛膜へと定着する。

4. Shhシグナル伝達制御とパラダイムシフト(V358A変異モデル)
脊椎動物の初期胚発生において、神経管のパターニング・四肢の形成・頭蓋顔面骨の発達など多岐にわたる形態形成をオーケストレートするのがヘッジホッグ(Hedgehog: Hh)シグナル伝達経路です。ARL13BはこのShhシグナル制御において、近年のゲノム編集研究によってこれまでの定説を根本から覆す驚くべき事実が明らかになりました。
💡 用語解説:ソニック・ヘッジホッグ(Shh)シグナル伝達とは
Shh(ソニック・ヘッジホッグ)は、胚発生において神経管の腹側パターニングや四肢の指の形成に必須の分泌性タンパク質(リガンド)です。ShhがPatched1(Ptch1)受容体に結合すると、Smoothened(Smo)というGタンパク質共役受容体様タンパク質が一次繊毛内に集積・活性化されます。活性化されたSmoは転写因子Gli(グリ)をアクチベーター型に変換し、核内で標的遺伝子の発現を制御します。哺乳類では、この一連のプロセスはすべて一次繊毛を舞台に展開されます。
従来のモデルと「Smoの繊毛内集積パラドックス」
従来の研究において、ARL13BはSmo(Smoothened)の繊毛内への移行と局在を制御する主要な因子であると考えられていました。ところが、ARL13Bを完全に欠失させた細胞では、Shhリガンドが全く存在しない状態であってもSmoが一次繊毛内に恒常的に異常集積するというパラドックスが生じます。にもかかわらず、Gliアクチベーターの産生は著しく減少し、下流へのシグナル伝達は完全に不全に陥ります。このパラドックスは長らく謎でしたが、V358A変異モデルの登場によって解決されました。
V358A変異モデルが覆したドグマ
研究者らはCRISPR/Cas9システムを用いてマウスの内因性Arl13b遺伝子座に点変異(V358A)を導入しました。この変異はARL13BのVxPxモチーフを特異的に破壊するものです。生成されたARL13B V358Aタンパク質は正常な発現レベルと、内在性・GAP依存性のGTP加水分解活性・ARL3に対するGEF活性といったすべての生化学的機能を完全に保持していました。しかし、逆行性輸送をブロックした条件下でも、この変異型タンパク質は一次繊毛内からは一切検出されませんでした。
驚愕すべきことに、一次繊毛の内部にARL13Bが全く存在しないこのArl13b V358A/V358Aホモ接合体マウスは、致死性を示す完全ノックアウトマウスとは対照的に、正常に生存し、完全な生殖能力を有し、発生過程におけるShhシグナル伝達も完全に正常に機能していました。
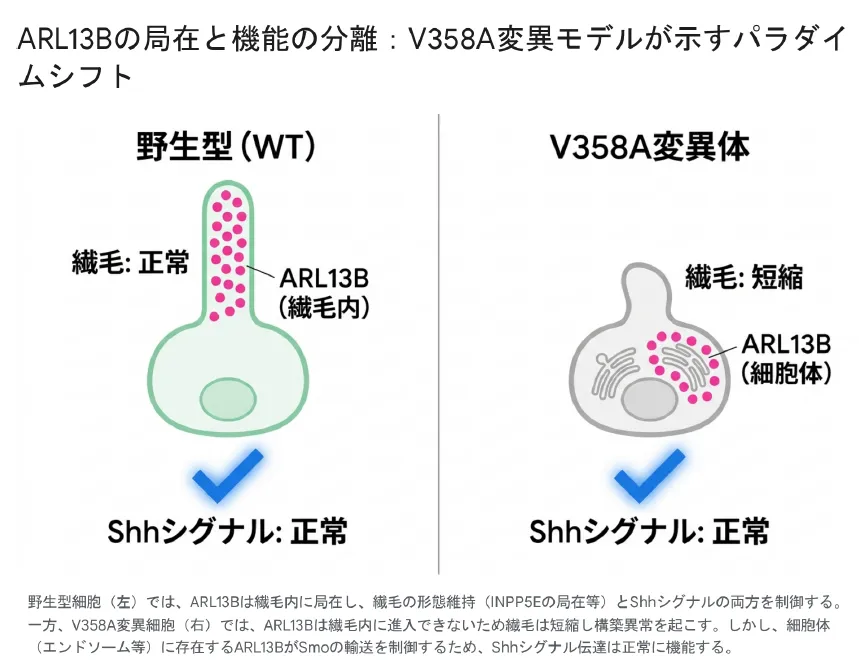
野生型細胞(左)ではARL13Bは繊毛内に局在し繊毛の形態維持とShhシグナルの両方を制御する。V358A変異細胞(右)ではARL13Bは繊毛内に進入できないため繊毛は短縮し構造異常を起こすが、細胞体(エンドソーム等)に存在するARL13BがSmoの輸送を制御するためShhシグナル伝達は正常に機能する。
🔑 V358A変異モデルがもたらした2つの決定的結論
①「構造と機能の解離(Uncoupling)」:ARL13Bが一次繊毛の長さを維持しARL3/INPP5Eを局在させる「繊毛構築機能」と、Shhシグナルカスケードを制御する「シグナル伝達機能」は、細胞内で完全に独立したプロセスとして分離可能です。
②「細胞体からの遠隔制御機構」:ARL13Bは繊毛内ではなく、細胞体(初期エンドソームや細胞膜上の背側ラッフルなど)の外部コンパートメントに存在することで、SmoをはじめとするShhコンポーネントのトラフィッキングを遠隔的に制御しています。
5. 臓器特異的な生理的機能:脳・網膜・精子
ARL13Bは全身の一次繊毛で遍在的に発現していますが、その機能的依存度や欠損時の表現型は、組織の特性や発生段階によって大きく異なります。とりわけ脳・網膜・精子という高度に特殊化した組織において、その機能は決定的な意味を持ちます。
大脳皮質の構築とニューロン移動制御
大脳皮質の緻密な層構造が形成されるためには、ARL13Bによる繊毛シグナリングの精緻な制御が不可欠です。ARL13Bは、大脳皮質構築の最初のステップである「極性化した放射状グリア(Radial glial)スキャフォールドの初期形成」に必須です。このスキャフォールドは、後に続くニューロンが正しい位置へ移動するための物理的なガイドとして機能します。さらに、分裂を終えた後有糸分裂介在ニューロンの長距離移動と皮質内での最終的な配置も直接的に制御しています。ARL13Bが喪失すると細胞骨格のリモデリングに重大な異常が生じ、皮質におけるニューロンの移動障害が引き起こされます。これらの機能欠陥は、ジュベール症候群における認知・運動機能障害の基礎的なメカニズムを説明するものです。
網膜視細胞の形態形成と視覚機能の維持
網膜に存在する光受容細胞(視細胞)の外節(Outer segment)は、ロドプシンなどの視覚タンパク質が高度に集積した膜ディスク構造で、進化的に一次繊毛が極度に特殊化・巨大化した形態です。ARL13Bは視細胞において、この外節にのみ特異的に局在します。
網膜特異的な条件付き欠損マウス(retArl13b⁻/⁻)を用いた解析により、生後6日目(P6)以降にARL13Bを欠損させると外節構造が全く形成されず、外節タンパク質群のトラフィッキングが完全に遮断されることが明らかになりました。生後10日目の超微形態解析では、基底小体が細胞膜へドッキングする初期プロセスは進行しているものの、成熟したトランジションゾーンや膜ディスク構造は一切形成されていませんでした。さらに成体マウスで後天的にARL13Bを欠損させても、微小管軸糸の短縮と外節の進行性退縮が確認されています。最終的には視覚応答の完全な喪失(失明)が引き起こされます。ジュベール症候群患者に高頻度で合併する重篤な網膜障害(レチノパシー)は、このARL13Bの視細胞における必須機能の喪失によって分子レベルで説明されます。
精子形成における不可欠な機能(2025年最新知見)
2025年に発表された最新の生殖生物学研究により、ARL13Bがオスの精子形成において他の体細胞組織とは全く異なる厳密なレベルで不可欠であることが判明しました。内因性ARL13Bを欠損したマウスに対して、機能ドメインを完全に保持した蛍光融合タンパク質「ARL13B-Cerulean」を発現させるレスキュー実験が行われ、このレスキューマウスは腎臓・脳・膵臓などの体細胞組織では一次繊毛の構造とShhシグナルが完全に回復し健康な成体へと成長しましたが、オスマウスは完全に不妊でした。
精子の重篤な欠陥
- 著しい精子減少症(Oligozoospermia)
- 前進運動能力の喪失
- 極端に短い鞭毛
- 非定型的な先体(Acrosome)構造
示唆される役割
V358Aマウス(ARL13Bが繊毛内に入れない)は生殖能力が正常に維持されます。このことから精子形成においてARL13Bは「細胞体(非繊毛コンパートメント)」における小胞輸送・核凝縮・先体形成の制御として機能していることが示唆されます。
精子の鞭毛はミトコンドリア鞘・外側粗大線維・線維鞘といった精子独自の巨大なアクセサリー構造体を備えており、これらを組み立てるためにはC末端に蛍光タンパク質が付加された人工的なARL13B-Ceruleanでは対応できない、内因性ARL13Bに固有の精密なタンパク質間相互作用が要求されることを示しています。
6. 関連疾患の病態メカニズム
ARL13Bの生化学的な機能不全または過剰発現は、広範なヒト疾患の病態に直接的に関与します。機能喪失型変異は重篤な先天性発生障害を引き起こす一方、成体組織での過剰発現や異常活性化は悪性腫瘍の進行を促進します。
ジュベール症候群(Joubert Syndrome)と繊毛病
💡 用語解説:ジュベール症候群(Joubert Syndrome: JS)とは
ジュベール症候群は、中脳-菱脳接合部の特異的な奇形(MRI画像で「大臼歯徴候:Molar tooth sign」が確認される)、顕著な発達遅滞・知的障害・筋緊張低下・呼吸窮迫・網膜症などを特徴とする稀な常染色体劣性(または一部X連鎖)遺伝性疾患です。一次繊毛の機能不全によって引き起こされる「繊毛病(シリオパシー)」の代表的疾患であり、現在30以上の原因遺伝子が同定されています。ARL13Bはその中の一つで、発見された変異は主にミスセンス変異です。
ARL13B遺伝子のミスセンス変異(患者から同定されたR77Q・R79Q・Y86C・R194C・R200Cなど)は、古典的なジュベール症候群の直接的な原因として同定されています。なお、ネフロン癆(Nephronophthisis)やメッケル症候群といった他の重篤な繊毛病コホートではARL13B変異は同定されておらず、変異の影響は古典的なJSの表現型に強く関連しています。
分子レベルの解析により、JS患者の変異(特にR79QやR200C)がARL13Bタンパク質に与える欠陥が詳細に解明されています。驚くべきことに、これらの変異はARL13B自身のGTP加水分解活性速度には有意な差をもたらしません(GTPアーゼとしての基本的な酵素能力は維持される)。その代わり、変異アミノ酸はARL3と相互作用する「GEFインターフェース」の構造的完全性を決定的に損ないます。
R77Q / R79Q 変異
生化学的影響:GTP加水分解活性は正常。しかしARL3とのGEFインターフェースが破壊され、ARL3の活性化速度が激減(R77Q変異で約1/13以下)。
臨床表現型:活性型ARL3の枯渇によりINPP5E等の繊毛局在が阻害され、大臼歯徴候・筋緊張低下・網膜症を発症。
R194C / R200C 変異
生化学的影響:C末端近傍の変異。GTPアーゼ活性は維持されるが、細胞内ARL3・GTPレベルの上昇が抑制されシグナルカスケードが遮断される。
臨床表現型:神経管のパターニング異常、大脳皮質のニューロン移動障害、重篤な視覚障害。
悪性腫瘍(グリオーマ・胃がん):ZEB1-ARL13B-VEGFR2軸
成体組織におけるARL13Bの過剰発現やシグナル異常は、悪性腫瘍の進行を強力に後押しする発癌性の機能を発揮します。特に胃がんや多形膠芽腫(GBM)と呼ばれる最も悪性度の高いグリオーマにおいて、ARL13Bの高発現は腫瘍サイズの増大・浸潤の深さ・患者の予後不良と極めて強い正の相関を示します。
胃がんにおけるメカニズム:ARL13BはSmo(Smoothened)のトラフィッキングと細胞内局在の安定性を高め、腫瘍細胞においてHedgehogシグナル伝達経路を持続的かつ過剰に活性化することで、無秩序な細胞増殖プログラムを駆動します。
💡 用語解説:ZEB1転写因子とは
ZEB1(Zinc finger E-box binding homeobox 1)は、上皮間葉移行(EMT)のマスターレギュレーターとして知られる転写因子です。グリオーマなどの悪性腫瘍において、ZEB1はARL13B遺伝子のプロモーター領域に直接結合し、ARL13Bの発現を転写レベルで強力に活性化することが明らかになっています。これがARL13Bの過剰発現を介した腫瘍増悪・血管新生促進という「ZEB1→ARL13B→VEGFR2軸」の起点となります。
グリオーマにおけるZEB1-ARL13B-VEGFR2血管新生軸:ZEB1→ARL13B転写活性化→Hedgehogシグナル活性化→VEGFA(血管内皮増殖因子A)の産生促進→周辺血管内皮細胞のVEGFR2への結合という連鎖が生じます。さらに腫瘍細胞から分泌されたARL13B自体が内皮細胞のVEGFR2の細胞内ドメインと直接相互作用し、VEGFA-VEGFR2シグナル伝達経路を相乗的に増幅させます。この増幅により内皮細胞の増殖・遊走・管腔形成が猛烈に促進され、腫瘍内への異常な血管ネットワークが構築されます。
動物モデルの実験では、GBMでARL13Bをノックダウンすると頭蓋内腫瘍の成長が顕著に遅延し、マウスの生存期間が延長することが確認されています。ZEB1-ARL13B-VEGFR2軸は、悪性固形腫瘍に対する新たな抗腫瘍血管新生療法の極めて有望な分子標的として注目されています。
増殖性硝子体網膜症(PVR)とTGF-β1/Smad3経路を介したEMT
💡 用語解説:上皮間葉移行(EMT)とは
上皮間葉移行(EMT:Epithelial-Mesenchymal Transition)とは、本来は上皮細胞として機能していた細胞が、不適切なシグナルに応答して間葉系細胞様の性質(高い遊走能・浸潤能・増殖能)へと脱分化するプロセスです。胚発生では正常なプロセスですが、成体では網膜疾患の線維化(PVR)やがんの転移に関与する病態生理学的に重要な現象です。
2025年の最新研究では、増殖性硝子体網膜症(PVR:網膜剥離の最も重篤な合併症)の進行においてARL13Bが重要な役割を持つことが明らかになりました。PVR患者の臨床検体(増殖膜)の解析では、間葉系マーカー(α-SMA)陽性の細胞領域でARL13Bの発現が著しく低下していることが確認されています。
インビトロでARL13Bをノックダウンすると、TGF-β1(PVR発生の最も強力な誘導因子)によって引き起こされる網膜色素上皮(RPE)細胞のEMTが劇的に促進されました。詳細な解析から、ARL13Bの喪失はSmad3の総タンパク質発現量を顕著に増加させ(転写レベルではなく翻訳後の安定化を介して)、TGF-β1刺激後わずか5分という初期段階からリン酸化活性型Smad3(pSmad3)を増大させます。この効果はSmad3を介したカノニカルなTGF-β1経路に特異的で、非カノニカルなERK経路やHedgehog経路への影響は見られませんでした。
これらの知見は、正常に機能するARL13B(および健全な一次繊毛構造)が、細胞が不必要に間葉系へと脱分化し過剰増殖するのを防ぐ「分子ブレーキ」として機能しており、その喪失が網膜組織の不可逆的な線維化とPVRの発症を直接駆動することを示しています。
7. 遺伝子検査について
ARL13B遺伝子の変異検索は、主にジュベール症候群(繊毛病)が疑われる場合に行われます。臨床的に大臼歯徴候(Molar tooth sign)、筋緊張低下、眼球運動異常、発達遅滞、網膜障害といった所見がある場合、繊毛病に関連する遺伝子を網羅的に解析できる遺伝子パネル検査や全エクソーム解析(WES)が推奨されます。
ジュベール症候群は現在30以上の原因遺伝子が知られており、表現型だけでは原因遺伝子を特定することが困難なため、両親と本人を含む「トリオ解析」が診断精度を高めるうえで有効です。ARL13B遺伝子変異が同定された場合、遺伝形式(常染色体劣性)を踏まえた遺伝カウンセリング、次子の出生前診断(羊水検査・絨毛検査)の検討、そして眼科・腎臓・神経科との集学的な管理体制の構築が重要となります。
よくある質問(FAQ)
🏥 繊毛病・遺伝性疾患の診断・遺伝カウンセリングについて
ジュベール症候群をはじめとする繊毛病・希少遺伝性疾患に関するご相談は、
臨床遺伝専門医が在籍するミネルバクリニックにお気軽にお問い合わせください。
関連記事
参考文献
- [1] Hanke-Gogokhia C, et al. The guanine nucleotide exchange factor Arf-like protein 13b is essential for assembly of the mouse photoreceptor transition zone and outer segment. J Biol Chem. 2017;292(51):21130-21143. [PubMed 29089384]
- [2] NCBI Gene: ARL13B ARF like GTPase 13B [Homo sapiens (human)]. Gene ID: 200894. [NCBI Gene]
- [3] Bharat V, et al. ARL13B regulates Sonic hedgehog signaling from outside primary cilia. eLife. 2020;9:e50434. [eLife]
- [4] Gotthardt K, et al. A G-protein activation cascade from Arl13B to Arl3 and implications for ciliary targeting of lipidated proteins. eLife. 2015;4:e11859. [eLife]
- [5] UniProt: ARL13B – ADP-ribosylation factor-like protein 13B [Homo sapiens]. UniProt ID: Q3SXY8. [UniProt]
- [6] Cantagrel V, et al. Mutations in the Cilia Gene ARL13B Lead to the Classical Form of Joubert Syndrome. Am J Hum Genet. 2008;83(2):170-179. [PMC2495072]
- [7] Wang H, et al. Arl13b promotes gastric tumorigenesis by regulating Smo trafficking and activation of the Hedgehog signaling pathway. Mol Oncol. 2017;11(8):1026-1044. [PMC5540784]
- [8] Zhou Y, et al. ARL13B promotes angiogenesis and glioma growth by activating VEGFA-VEGFR2 signaling. J Exp Clin Cancer Res. 2022;41(1):318. [PMC10158193]
- [9] Loss of a primary cilia protein ARL13B promotes TGFβ-1 induced EMT in proliferative vitreoretinopathy. Front Cell Dev Biol. 2025;13:1661658. [Frontiers]
- [10] ARL13B-Cerulean rescues Arl13b-null mouse from embryonic lethality but fails to rescue male fertility. bioRxiv. 2025. [bioRxiv]
- [11] Larkins CE, et al. Arl13b regulates ciliogenesis and the dynamic localization of Shh signaling proteins. Mol Biol Cell. 2011;22(23):4694-4703. [PMC3226485]
- [12] Miertzschke M, et al. Biochemical characterization of purified mammalian ARL13B protein indicates that it is an atypical GTPase and ARL3 guanine nucleotide exchange factor (GEF). J Biol Chem. 2017;292(26):11091-11108. [PubMed 28487361]
- [13] Mariani LE, et al. The G-domain of the Joubert syndrome protein ARL13B interacts with tubulin and contributes to uniform distribution along the cilium. Invest Ophthalmol Vis Sci. 2016;57(13):5533-5545. [IOVS]
- [14] Ying G, et al. Ciliary ARL13B Is Essential for Vision and Morphogenesis of Cone Outer Segments. J Neurosci. 2025;45(34):e0752252025. [J Neurosci]
- [15] Gotthardt K, et al. A G-protein activation cascade from Arl13B to Arl3 and implications for ciliary targeting of lipidated proteins. eLife. 2015. [PMC4868535]






