承認済シンボル:DYNC2H1
遺伝子名:dynein cytoplasmic 2 heavy chain 1
参照:
HGNC: 2962
AllianceGenome : HGNC : 2962
NCBI:79659
Ensembl :ENSG00000187240
UCSC : DYNC2H1 (ENST00000375735.7) from GENCODE V47
遺伝子OMIM番号603297
●遺伝子のlocus type :タンパク質をコードする
●遺伝子のグループ:
Dynein 2 complex subunits
●遺伝子座: 11q22.3
●ゲノム座標: 11:103,109,426-103,479,863
遺伝子の別名
DHC2
DYH1B
dynein, cytoplasmic 2, heavy chain 1
hdhc11
遺伝子の概要
DYNC2H1遺伝子は、ダイニン-2複合体と呼ばれるタンパク質の一部を生成するための指示を提供します。ダイニン-2は、細胞の繊毛内に存在し、繊毛の形成および維持に不可欠な役割を果たしています。繊毛は、細胞の表面に存在する毛のような小さな構造で、細胞の移動や信号伝達に重要です。
IFT(繊毛内輸送)は、繊毛内で必要なタンパク質や分子を運搬するプロセスであり、繊毛の構造維持と機能に欠かせません。ダイニン-2は、このIFTの逆行性輸送(細胞の先端から基底に向かう輸送)に関与しており、これにより繊毛の健康を保つことができます。
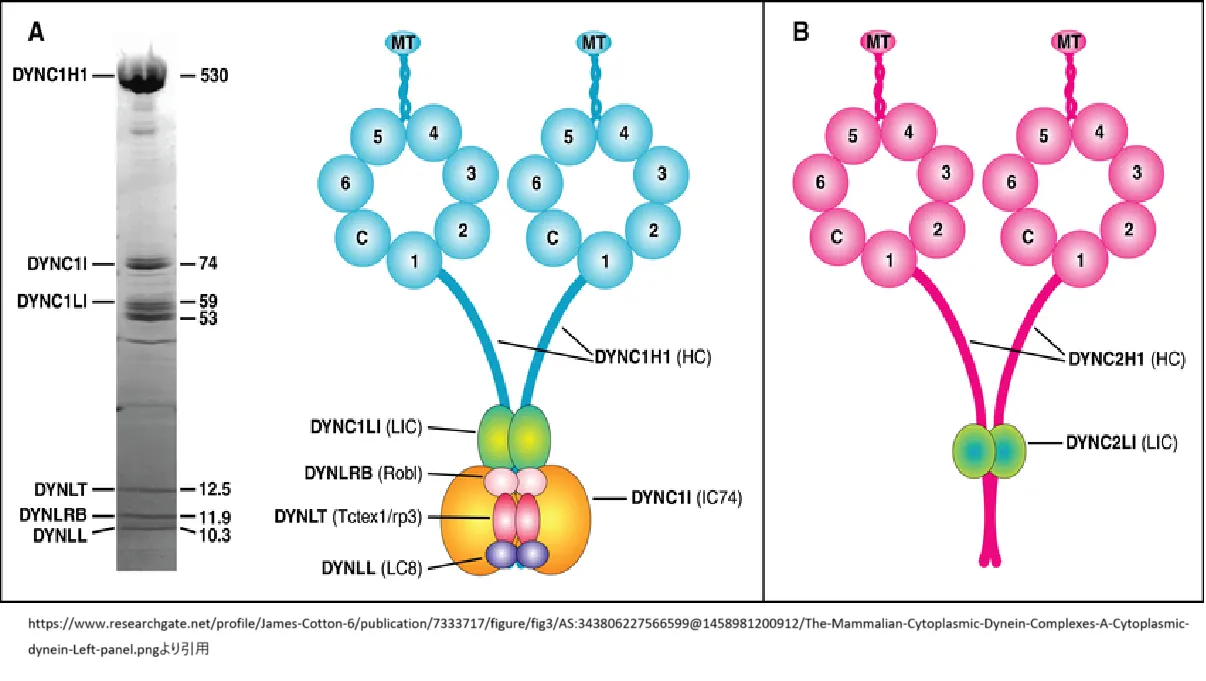
https://www.researchgate.net/profile/James-Cotton-6/publication/7333717/figure/fig3/AS:343806227566599@1458981200912/The-Mammalian-Cytoplasmic-Dynein-Complexes-A-Cytoplasmic-dynein-Left-panel.png
より引用
真核細胞質ダイニン複合体(A)細胞質ダイニン。(左のパネル)免疫親和性精製ラット脳細胞質ダイニンのポリペプチド。 ゲルの右側にポリペプチドの分子量(kDa)が示され、左側にファミリー名のコンセンサスが示されています。(右のパネル)細胞質ダイニン複合体のサブユニットの結合構造モデル。細胞質ダイニン複合体のコアは、N末端領域を介してホモダイマーを形成する2つのDYNC1H1重鎖から構成されています。モータードメインは重鎖のC末端にあり、中央の空洞を取り囲む7つの密度のリングから構成される、約350kDaの大きな球状頭部です。6つの密度はAAAドメイン(1~6)です。AAAドメイン1はATP加水分解の部位です。微小管結合ドメインは、リングの反対側にあるAAAドメイン4と5の間に見られる突起です。Cは、7番目の密度を形成する重鎖のC末端です。2つのDYNC1I中間鎖(IC74)とDYNC1LI軽鎖中間鎖は、重鎖のN末端の重複領域で結合し、重鎖の二量体化ドメインと重複します。3つの軽鎖ファミリー、DYNLT、Tctex1軽鎖、DYNLRB、Roadblock軽鎖、およびDYNLL、LC8軽鎖の二量体は、中間鎖二量体に結合します。(B) 細胞質ダイニン2複合体、サブユニット結合の構造モデル。このダイニン複合体は IFT において独特な役割を果たしており、IFT ダイニンと呼ばれることもあります。構造予測によると、重鎖 DYNC2H1 は細胞質ダイニンおよび軸糸ダイニンと類似しています。この複合体のサブユニットとして知られているのは、33~47kDaのポリペプチドであるDYNC2LI1のみであり、これは細胞質ダイニン軽鎖と関連しています。中間鎖や軽鎖はまだ同定されていません。
遺伝子と関係のある疾患
遺伝子ファミリー
1. 軸糸ダイニンは、線毛や鞭毛の運動を制御し、これらの構造内で微小管の局所的な滑り運動を引き起こすことで、細胞小器官の振動運動を支えます。
2. 細胞質ダイニンは、細胞内での物質輸送を含むさまざまな活動に関与しており、細胞小器官の移動や、シグナル伝達、分裂紡錘体の形成などを助けます。
ダイニンは重鎖、軽中間鎖、中間鎖、軽鎖から構成される多サブユニットタンパク質です。重鎖(分子量500kD以上)は、実際のモーター機能を担っており、ATPの加水分解によってエネルギーを供給し、運動を引き起こします。中間鎖は、ダイニン分子を適切な細胞内標的に誘導する役割を果たし、軽中間鎖は主に細胞質ダイニンに特有で、ATPアーゼの要素を持っています。また、8~25kDの軽鎖も複合体に含まれ、全体の構造と機能に寄与しています【Gibbons et al., 1994】【Vaughan et al., 1996】。
遺伝子の発現とクローニング
その後、Vaisbergら(1996年)は、RT-PCR法を用いて、ダイニン重鎖の保存領域に基づくプライマーからDHC2をコードするcDNAを単離しました。DHC2は、ヒトDYH1bの相同体とされ、主にゴルジ装置に局在することが確認されました。この結果から、DHC2が正常なゴルジ体の形成に関与することが示唆されます。また、DHC2は線毛や鞭毛を持たない組織でも発現しており、ゴルジ体の機能に関連していると考えられます【Vaisbergら、1996】。
Dagoneauら(2009年)は、ヒトDYNC2H1遺伝子が4,314アミノ酸からなるタンパク質をコードしていることを明らかにしました。このタンパク質は、逆行性繊毛輸送を担う重要な役割を果たしており、細胞内の微小管に沿った物質の輸送に不可欠です【Dagoneauら、2009】。
生化学的特徴
マッピング
遺伝子の構造
分子遺伝学
Dagoneau ら(2009年)の研究では、DYNC2H1遺伝子における二アレル性突然変異が、SRTD3の患者に確認されました。彼らはこの研究結果に基づき、ATD3とSRPS IIIは同じスペクトラムに属するアレリック疾患であると結論付けました。この研究は、これらの疾患が遺伝的に関連していることを示す重要な証拠となりました。
Schmidts ら(2013年)は、SNPマッピング、エクソームシークエンシング、サンガーシークエンシングを組み合わせた研究を通じて、57家族から71人のSRTD3患者を調査しました。その結果、DYNC2H1遺伝子において34の新しい変異が発見されました。これらの変異には、13の終止変異と21のミスセンス変異が含まれており、一部は遺伝子全体に広がっていました。興味深いことに、2つの終止変異を持つ患者はいませんでした。このことから、DYNC2H1の変異は完全な機能喪失ではなく、部分的な機能低下(低形成)を引き起こすと考えられます。また、いくつかの患者の細胞を用いた研究により、線毛の逆行輸送(IFT)に欠陥があり、線毛先端にタンパク質が異常に蓄積することが確認されました。
一方で、Merrill ら(2009年)は、近親婚の家族に属するSRTD3患者と、短肋多指症候群III型(SRPS3)と診断された孤立例の患者において、DYNC2H1遺伝子にホモ接合または複合ヘテロ接合の変異を発見しました。彼らの研究は、DYNC2H1が軟骨形成における線毛機能に重要な役割を果たしていることを示唆しています。
また、Thiel ら(2011年)の研究では、短肋多指症II型患者において、DYNC2H1遺伝子とNEK1遺伝子にそれぞれ1つのミスセンス変異が見つかりましたが、両遺伝子の変異は単独では疾患を引き起こさないとされています。さらに、El Hokayem ら(2012年)は、8人の短肋多指症II型患者の中でDYNC2H1の複合ヘテロ接合変異を4例発見しました。
最後に、Badiner ら(2017年)は、重篤な短肋多指症患者3人においてDYNC2H1遺伝子の複合ヘテロ接合変異を発見し、これには保存された残基におけるミスセンス変異が含まれていました。これらの変異は線毛形成と骨格異常の発生に重要な影響を与えていると考えられています。これらの研究は、DYNC2H1遺伝子がSRTD3および関連疾患における中心的な役割を果たしていることを示しています。
動物モデル
この研究では、線毛順行輸送タンパク質Ift172の量を野生型の約60%に減少させると、Dync2h1 -/-マウスにおける異常な繊毛形態やヘッジホッグシグナル伝達の欠損が部分的に回復し、胚の生存期間が延長されることが観察されました。さらに、Ift122を減少させることも同様に、Dync2h1 -/-の表現型を抑制する効果がありました。
これらの結果を基に、Ocbina ら(2011)は、Dync2h1が正常な線毛構造を維持し、繊毛軸に沿ったタンパク質の逆行輸送に必要であると結論づけています。この研究は、Dync2h1が繊毛の機能およびヘッジホッグ経路における重要な役割を果たしていることを示す重要な証拠を提供しました。
アレリックバリアント
DYNC2H1, MET1991LEU
2人の子供が絞扼性胸部異形成症(SRTD3; 613091)を発症した近親結婚のモロッコ人家族において、Dagoneau ら(2009年)は、DYNC2H1 遺伝子における2つのミスセンス変異のホモ接合性を特定しました。1つは、ヌクレオチド5971におけるA-to-T転換で 。これはコドン1991におけるメチオニンからロイシンの置換(M1991L)を招き、ヌクレオチド11284におけるAからGへの転位は、コドン3762におけるメチオニンからバリンの置換(M3762V;603297.0002)を招きます。これらの変異のいずれかがシスで1つのアレリックのみに存在するキャリアは、無症候性でした。
0.0002 多指症を伴わない短肋骨胸郭異形成症3型
DYNC2H1、MET3762VAL
Dagoneau ら(2009年)により、無気肺性胸郭異形成症(SRTD3; 613091)患者において複合ヘテロ接合体の状態で発見された DYNC2H1 遺伝子の met3762-to-val(M3762V)変異に関する考察については、603297.0001 を参照してください。
.0003 多指症を伴わない短肋骨胸郭異形成症3型
DYNC2H1、29-BP INS、NT654
非近親のフランス人家族で、臨床的に無呼吸性胸郭異形成症(SRTD3; 613091)と診断された2人の胎児がいた。Dagoneau ら(2009)は、 DYNC2H1遺伝子のエクソン5におけるフレームシフト変異、654番目の位置に続く29ヌクレオチドの挿入、およびミスセンス変異(603297.0004)が複合ヘテロ接合であることを確認しました。この挿入変異により、コドン219でグルタミン酸からロイシンへの置換が起こり、その2アミノ酸後に終止コドンが現れるフレームシフトが起こりました(Glu219LeufsTer2)。
0.0004 短肋骨胸郭異形成症3 多指症なし
DYNC2H1、ASP3015GLY
非近親のフランス人家族で、臨床的に無呼吸性胸郭異形成症(SRTD3; 613091)と診断された2人の胎児について、Dagoneau ら(2009)は、 DYNC2H1遺伝子のエクソン57のヌクレオチド9044におけるA-to-G転位が、コドン3015(D3015G)におけるアスパラギンからグリシンへの置換を引き起こしていることを確認しました。もう一方のアレリック(603297.0003)にはフレームシフト変異が認められました。
SRTD3を発症したオランダの3家族(JATD-1、JATD-2、JATD-6)の無関係な患者において、Schmidtsらは(2013年)、DYNC2H1遺伝子における他の変異との複合ヘテロ接合状態でD3015G変異を特定しました。
0.0005 多指症を伴わない短肋骨胸郭異形成症3型
DYNC2H1、ILE1240THR
非近親婚のフランス人同士の間に生まれた、臨床診断で閉塞性胸部異形成症(SRTD3; 613091)と診断された19歳の患者について、Dagoneau ら(2009年)は、DYNC2H1遺伝子における2つの変異について複合ヘテロ接合性を特定しました。 エクソン25のヌクレオチド3719におけるTからCへの転移、その結果コドン1240におけるイレからスレへの置換(I1240T)、およびエクソン66のヌクレオチド10063におけるGからTへの転換、その結果グリ3355からテル(G3355X)への置換(603297.0006)が確認されました。
SRTD3を持つドイツ人女性(JATD-8)において、Schmidts ら(2013年)は、DYNC2H1遺伝子における別の変異との複合ヘテロ接合状態でI1240T変異を特定しました。
0.0006 短肋骨胸郭異形成症3型(多指症なし)
DYNC2H1、グリシン3355番目のテール変異
Dagoneau ら(2009 年)により、無酸素性胸部異形成症(SRTD3; 613091)患者において複合ヘテロ接合体の状態で発見された DYNC2H1 遺伝子のグリシン3355-テロメア(G3355X)変異に関する考察については、603297.0005 を参照してください。
.0007 多指症を伴うまたは伴わない短肋骨胸郭異形成症3
DYNC2H1, ARG587CYS
4人の患児は、臨床的に肋骨短縮多指症候群III型(SRTD3;613091)と診断された従兄弟同士の親を持つ(家族R01-314)。Merrillら(20 2009)は、DYNC2H1遺伝子のエクソン12のヌクレオチド1759におけるC-to-T転位のホモ接合性を検出し、これはアルギニン587がシステインに置換するアミノ酸置換(R587C)を引き起こすことが予測されました。 発症していない両親と兄弟は、この突然変異に対してヘテロ接合性でした。著者らは、この家族では表現型の多様性が認められると指摘しています。女性発端者には多趾症は認められなかったのに対し、他の3人の兄弟姉妹には両手および両足の軸後性多趾症が認められたからです。
0.0008 短肋骨胸郭異形成症3型および多趾症
DYNC2H1、ARG2205HIS
非血縁家族から臨床的に短肋多指症候群III型(SRTD3; 613091)と診断された患者において、Merrill ら(2009年)は、DYNC2H1遺伝子のエクソン41におけるヘテロ接合型6614G-A転位を発見しました。これにより、コドン2205(R2205H)におけるアルギニンがヒスチジンに置換されました。もう一方のDYNC2H1アレリックにはナンセンス変異が認められました(603297.0009)。
0009 多指症を伴う短肋骨胸郭異形成症3
DYNC2H1, ARG2838TER
非血縁家族から臨床的に短肋骨多指症候群III型(SRTD3; 613091)と診断された患者において、Merrill ら(2009年)は DYNC2H1遺伝子のエクソン53における8512C-T転位が早期タンパク質終結(R2838X)を引き起こし、ミスセンス変異(603297.0008)との複合ヘテロ接合性が認められました。
短肋骨胸郭異形成症3型と多指症
DYNC2H1, 2-BP 欠失/挿入
非血縁家族から臨床診断で短肋多指症候群III型(SRTD3; 613091)と診断された患者において、Merrill ら(2009年)は DYNC2H1 遺伝子における複合ヘテロ接合性変異を検出しました。一方のアレリックでは、エクソン5の2塩基連続の変化(624_625GT-AA)が認められました。最初のヌクレオチド変化は、コードされたアミノ酸を変化させることなくコドン208の最後の塩基を変化させ、2番目の変化はフェニルアラニン209からイソロイシン(F209I)への置換を予測しました。もう一方のアレリックでは、スプライス供与部位の変異(603297.0011)が認められました。
.0011 多指症を伴う短肋骨胸郭異形成症 3
DYNC2H1、IVS33、G-T、+1
臨床診断で短肋多指症候群III型(SRTD3; 613091)と診断された患者において、Merrill ら(2009)は、DYNC2H1 遺伝子のイントロン33のスプライス供与部位の変異(IVS33+1G-T)のヘテロ接合性を発見しました。逆転写酵素および定量的 PCR により、その結果生じる転写産物がナンセンス媒介性分解を受けることが示されました。もう一方のアレルのエクソン 5 には 2 塩基対の置換が認められました(603297.0010)。
0012 多指症を伴う短肋骨胸郭異形成症3
DYNC2H1, GLN1537ARG
非血縁関係にあるフランス人家族から、臨床診断で短肋骨多指症候群3型(SRTD3; 613091)と診断された胎児について、Dagoneau ら(2009)は DYNC2H1遺伝子のエクソン30のヌクレオチド4610におけるA-to-G転位が、コドン1537(Q1537R)におけるグルタミンからアルギニンへの置換を引き起こしていることが確認されました。この突然変異は、別のミスセンス変異(G2461V; 603297.0013)との複合ヘテロ接合性で発見されました。
0013 多指症を伴う短肋骨胸郭異形成症3
DYNC2H1, GLY2461VAL
非血縁関係にあるフランス人家族から、臨床診断で短肋骨多指症候群3型(SRTD3; 613091)と診断された胎児について、Dagoneau ら(2009)は DYNC2H1遺伝子のエクソン45のヌクレオチド7382におけるG-to-T転換が同定され、その結果、コドン2461(G2461V)でグリシンがバリンに置換されました。この変異は、別のミスセンス変異(Q1537R; 603297.0012)との複合ヘテロ接合性で発見されました。
0014 多指症を伴わない短肋骨胸郭異形成症3型
DYNC2H1, THR1987ALA
マダガスカル出身の非血縁夫婦の子供で、臨床診断で短肋多指症候群III型(SRTD3; 613091)と診断された3人の胎児について、Dagoneau ら(2009年)は、DYNC2H1遺伝子における複合ヘテロ接合性の変異を特定しました。父方のアレリックにはエクソン38における5959A-G転位があり、T1987A置換が起こりました。また、母方のアレリックには エクソン67における1bp欠失(10130delT;603297.0015)により、フレームシフトと早期終結(Leu3377CysfsTer34)が起こりました。
0015 短肋骨胸郭異形成症3型(多指症なし)
DYNC2H1、1-BP欠失、10130T
Dagoneau ら(2009 年)により、III 型短肋多指症候群(SRTD3; 613091)の 3 人の胎児で複合ヘテロ接合型として発見された DYNC2H1 遺伝子における 1bp 欠失(10130delT)に関する考察については、603297.0014 を参照してください。
.0016 短肋骨胸郭異形成症 3/6 多指症、二遺伝子
DYNC2H1、GLY3916ASP
非血縁のドイツ系家族から臨床診断で短肋多指症候群III型(SRTD3; 613091)と診断された患者において、Thiel ら(2011年)は、DYNC2H1遺伝子におけるヘテロ接合性11747G-A転位を特定し、 DYNC2H1遺伝子におけるグリシン3916からアスパラギン酸への置換(G3916D)をもたらすヘテロ接合型11747G-A変異を同定しました。この個体は、NEK1遺伝子における1bp挿入(1640insA)のヘテロ接合型(604588.0003)でもありました。いずれの遺伝子にも2番目の突然変異は認められず、両親はそれぞれ1つの突然変異のヘテロ接合型であり、382人の集団に一致する対照染色体のいずれにも認められませんでした。したがって、2アレル性複合遺伝が示唆されました。
0.0017 短肋骨胸郭異形成症3型および多指症
DYNC2H1、ARG2662GLN
妊娠15週で中絶された短肋多指症候群III型(SRTD3; 613091)と臨床診断された男性胎児において、El Hokayem ら(2012年)は、DYNC2H1遺伝子における2つのミスセンス変異による複合ヘテロ接合性を特定しました。エクソン49における7985G-A転位は、アルギニン2662がグルタミンに置換する(R2662Q)結果となり、エクソン46における7486C-T転位は、ATP結合および加水分解ドメインにおけるプロリン2496がセリンに置換する(P2496S;603297.0018)結果となります。非近親交配のベトナム人の両親は、それぞれがこれらの突然変異の1つに対してヘテロ接合性であり、いずれの突然変異も200人の対照者の染色体には認められませんでした。.
0018 短肋骨胸郭異形成症3型および多指症
DYNC2H1、PRO2496SER
El Hokayem ら(2012年)により、III型短肋多指症(SRTD3; 613091)の胎児で複合ヘテロ接合体の状態で発見されたDYNC2H1遺伝子のpro2496-to-ser(P2496S)突然変異に関する考察については、603297.0017を参照してください。
0019 短肋骨胸郭異形成症3型多指症
DYNC2H1、ARG330CYS
15週で中絶された妊娠から、短肋骨多指症候群3型(SRTD3;613091)と臨床診断された男性胎児について、El Hokayem ら(2012年)は、 DYNC2H1遺伝子におけるミスセンス変異とフレームシフト変異の複合ヘテロ接合性を特定しました。エクソン6における988C-T転位により、N末端領域でアルギニン330がシステインに置換(R330C) 1、エクソン53における1bpの欠失(8534delA)により、早期終止コドン(Asn2845IlefsTer8;603297.0020)が生じると予測されるフレームシフトが起こります。非近親交配のハイチ人の両親は、それぞれがこれらの突然変異の1つに対してヘテロ接合であり、200の対照染色体にはいずれも認められませんでした。
0020 短肋骨胸郭異形成症3型および多指症
DYNC2H1、1-bp欠失、8534A
短肋多指症候群3型(SRTD3; 613091)の胎児で複合ヘテロ接合状態で発見されたDYNC2H1遺伝子(8534delA)の1bp欠失(603297.0019)については、El Hokayemら(2012年)による考察を参照してください。







