承認済シンボル:CYP27B1
遺伝子名:cytochrome P450 family 27 subfamily B member 1
参照:
HGNC: 2606
NCBI:1594
遺伝子OMIM番号
Ensembl :ENSG00000111012
UCSC : uc001spz.2
AllianceGenome : HGNC : 2606
遺伝子のlocus type :タンパク質をコードする
遺伝子のグループ:Cytochrome P450 family 27
遺伝子座: 12q14.1
●ゲノム座標: (GRCh38): 12:57,762,334-57,767,078
遺伝子の別名
25 hydroxyvitamin D3-1-alpha hydroxylase
25-hydroxyvitamin D-1 alpha hydroxylase, mitochondrial
25-OHD-1 alpha-hydroxylase
CYP1alpha
CYP27B
cytochrome p450 27B1
cytochrome P450 subfamily XXVIIB polypeptide 1
cytochrome P450, family 27, subfamily B, polypeptide 1
cytochrome P450C1 alpha
cytochrome P450VD1-alpha
P450c1
VD3 1A hydroxylase
遺伝子の概要
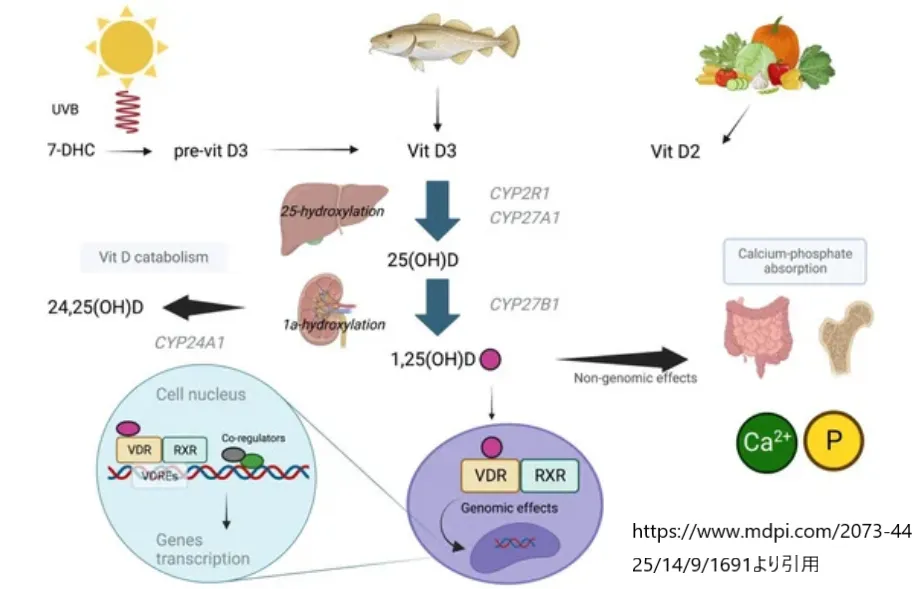
https://www.mdpi.com/2073-4425/14/9/1691
より引用
活性化されたビタミンD、すなわちカルシトリオールは、骨や歯の健康維持に必要なミネラル、特にカルシウムとリン酸の吸収、代謝、およびバランスの維持に重要な役割を果たします。具体的には、腸からのカルシウムとリン酸の吸収を促進し、それによって骨の形成と維持に必要なミネラルの供給を支援します。
さらに、ビタミンDは骨や歯の健康に限らず、免疫システムの機能、細胞成長、炎症の軽減など、体内の他の多くの生理的プロセスにも関与しています。ビタミンDの不足は、骨軟化症や骨粗鬆症などの骨疾患のリスクを高めるだけでなく、免疫系の機能低下や特定の慢性病のリスク増加にも関連しています。したがって、CYP27B1遺伝子およびそれによってコードされる1α-水酸化酵素の機能は、ビタミンDの効果的な活性化とその多岐にわたる生理的役割の実現に不可欠です。
紫外線によって人の表皮で合成されるビタミンD3(コレカルシフェロール)と、植物で生成される食事性ビタミンD2(エルゴカルシフェロール)は、そのままの形では生物学的に活性がありません。これらのビタミンDは、体内で複数の代謝ステップを経て活性形に変換される必要があります。ビタミンDの活性形である1-α,25-ジヒドロキシビタミンD3(1,25(OH)2D3、またはカルシトリオール)は、生物学的な効果を発揮する主要な形態です。
CYP27B1遺伝子は、この重要な過程である25-ヒドロキシビタミンD3から1,25(OH)2D3への変換を触媒する酵素、25-ヒドロキシビタミンD3-1-α-水酸化酵素(1-α-(OH)ase)をコードしています。この酵素は主に腎臓の近位尿細管で発現し、ビタミンDの最終活性代謝物である1,25(OH)2D3の生成を促進します。
1,25(OH)2D3は核内ビタミンD受容体(VDR)に結合して活性化し、その結果、カルシウムとリンのホメオスタシスの維持、骨形成と維持、細胞の分化と増殖、免疫応答の調節など、多様な生理的プロセスを制御します。VDRと1,25(OH)2D3の相互作用は、遺伝子発現の調節を介してこれらのプロセスに影響を及ぼし、健康な骨格システムの維持だけでなく、多くの慢性疾患の予防にも寄与します。
Takeyamaら(1997)、Liberman and Marx(2001)、Koren(2006)による要約は、ビタミンDの代謝とその生理的重要性に関する理解を深め、ビタミンDが人体の健康維持に不可欠であることを示しています。CYP27B1遺伝子やVDRの変異はビタミンDの代謝異常や感受性疾患のリスクを高める可能性があり、これらの遺伝子の研究は将来の医療戦略の開発に貢献することが期待されています。
遺伝子の発現とクローニング
一方、Fuら(1997年)は、P450c1-αの発現が誘導されるヒトケラチノサイト由来のP450c1-α cDNAをクローニングし、塩基配列を決定しました。ノーザンブロット解析では、ケラチノサイトで2.5kbの転写産物が検出され、RT-PCRによってヒトの腎臓、脳、精巣での発現が確認されました。このタンパク質は、ミトコンドリアのシトクロムP450酵素に類似したトポロジーを持つことが予測され、1-α-水酸化酵素活性を示しました。また、サザンブロット法によりヒトゲノムにはP450c1-α遺伝子が1コピーしか存在しないことが示されました。
St-ArnaudらおよびMonkawaらによってもヒト1-α-ヒドロキシラーゼ遺伝子がクローニングされ、Kitanakaら(1998年)はヒト腎臓cDNAライブラリーから同遺伝子を単離し、マウスタンパク質と82%の相同性を持つことを発見しました。彼らの研究では、腎組織でのみ2.4kbの1-α-水酸化酵素mRNA転写物が同定されました。
Diazら(2000年)は、培養ヒト滑膜栄養細胞と胎盤細胞における25-ヒドロキシビタミンD3-1-α-ヒドロキシラーゼ遺伝子発現の存在を確認しました。これは、ヒト胎盤での1-α-(OH)aseの存在と絨毛細胞における25-(OH)D3の1,25-ジヒドロキシビタミンD3への変換が酵素的な1-α-水酸化反応によるものであることを示唆しています。
しかし、Vanhookeら(2006年)の研究では、マウスの皮膚や初代ケラチノサイトではCyp27b1の発現を示す証拠が見つかりませんでしたが、腎臓と胎盤では発現が確認されました。
これらの研究結果は、1-α-ヒドロキシラーゼ遺伝子とそのタンパク質の機能および発現に関する重要な洞察を提供しており、ビタミンD代謝とその生理的役割の理解を深めています。
遺伝子の構造
Kitanakaら(1998年)は、ヒトCYP27B1遺伝子が約4.8kbの領域に9つのエキソンを含むことを決定しました。
Kongら(1999年)の研究では、1-α-OHase遺伝子が9個のエキソンを含み、約6.5kbと1.4kbの5-プライムフランキング領域にまたがることが明らかにされました。プロモーター解析により、PTHと1,25(OH)2D3によるこの遺伝子の発現制御の生理学的基盤に関する洞察が得られました。
Turunenら(2007年)は、CYP27B1プロモーター近位の転写開始部位の2.6kbおよび3.2kb上流にビタミンD応答エレメント(VDRE)を同定しました。これらの上流エレメントはMETTL1遺伝子のイントロン内に位置しています。
これらの研究は、ヒトCYP27B1遺伝子の遺伝子構造とその制御機構に関する貴重な情報を提供しており、ビタミンD代謝経路におけるこの遺伝子の役割の理解を深めています。
マッピング
Fuらの研究では、体細胞ハイブリッド解析を用いて、P450c1-α遺伝子がビタミンD依存性くる病タイプ1A(VDDR1A;264700)がマッピングされた12番染色体に位置していることを明らかにしました。この遺伝子は、ビタミンDの代謝経路において重要な役割を担うことが知られています。
一方、St-Arnaudらの研究では、蛍光in situハイブリダイゼーション(FISH)を用いて、CYP27B1遺伝子を12q13.1-q13.3にマッピングしました。CYP27B1遺伝子は、ビタミンDをその活性形である1,25-ジヒドロキシビタミンD3に変換する酵素、1α-ヒドロキシラーゼをコードしています。この活性形のビタミンDは、カルシウムとリンの代謝に不可欠であり、骨の健康に重要な役割を果たします。
これらの遺伝子のマッピングは、ビタミンD依存性くる病や他のビタミンD関連疾患の病態生理学を理解するうえで重要なステップです。また、これらの遺伝子の正確な染色体位置の特定は、これらの遺伝子の変異が関与する疾患の診断や治療において、将来的に役立つ可能性があります。ビタミンD代謝経路の遺伝子に対する詳細な理解は、これらの疾患の予防や管理に新たなアプローチをもたらすかもしれません。
遺伝子の機能
25(OH)Dの1-α位での水酸化作用は腎尿細管細胞に限らず、ヒト胎盤の十二指腸細胞もカルシトリオールを生成し、この酵素活性はフィードバック機構で制御されることが示されています。Glorieuxらによる1995年の研究では、ビタミンD依存性くる病1A型の患者の胎盤十二指腸細胞からこの酵素活性が欠如していることが発見され、十二指腸酵素と腎酵素が同じ遺伝子によってコードされている可能性が示唆されました。
Takeyamaらの1997年の研究では、ビタミンDレセプターを欠損したマウスで1-α-ヒドロキシラーゼの発現が低下しており、この酵素の活性は正常マウスではホルモン活性型ビタミンDによって抑制されるが、VDR欠損マウスでは抑制されないことが示されました。これは、活性型ビタミンD合成の負のフィードバック制御がリガンド結合したVDRを介して行われることを示唆しています。
Brenzaらの1998年の研究では、マウスの1-α-水酸化酵素遺伝子のプロモーター活性とその制御機構が明らかにされ、PTHがこのプロモーターをcAMPを介して活性化することが示されました。MurayamaらとShinkiらの研究も、CYP27B1遺伝子の制御が転写レベルで行われていることを裏付けています。
Zehnderらの2001年の研究は、1-α-水酸化酵素の腎外での分布を明らかにし、さまざまな組織での特異的な役割を示唆しています。
山本らによる2005年の研究では、さまざまな点変異を持つマウスのCyp27b1遺伝子を用いて実験を行い、ヒトのCYP27B1遺伝子のThr409に相当するマウスのSer408が、25(OH)D3(ビタミンDの一形態)の25-水酸基と相互作用し、その後の1α水酸化反応における水分子の除去に不可欠であることを明らかにしました。また、Gln65も基質結合に関与していることが示されましたが、この部位の変異は、ヘムを含まないCyp27b1のアポタンパク質の形成を引き起こすことがわかり、タンパク質の構造形成に重要な役割を果たしていることが示唆されました。
Liuらの2006年の研究では、マイコバクテリアのリガンドがTLR2とTLR1を活性化させ、この活性化が単球とマクロファージにおけるVDRとCYP27B1の発現を増加させることが明らかにされました。ビタミンDで処理した単球では、CYP24(ビタミンD 24-水酸化酵素)とカテリシジン(抗菌ペプチド)の発現が増加しましたが、DEFB4の発現は変わりませんでした。ビタミンD処理により、結核菌に感染したマクロファージ内の生菌数が減少しました。さらに、アフリカ系アメリカ人の血清にビタミンDを補充することで、CAMP誘導の欠損が逆転することが示されました。
Schauberらによる2007年の研究では、ヒトの皮膚創傷がTLR2、CD14、CYP24A1の発現を上昇させることが確認されました。ビタミンD3は、ケラチノサイトにおけるTLR2とCD14の発現を増強し、CYP27B1の発現を増加させることで、1,25D3応答性遺伝子の発現を促進しました。これにより、ビタミンD3が自然免疫において重要な役割を果たしていることが示唆されました。
Turunenらの2007年の研究では、1,25(OH)2D3(活性型ビタミンD3)のレベルが産物阻害によって制御されており、CYP27B1の発現がVDRを介して1,25(OH)2D3によってダウンレギュレートされることが確認されました。この研究では、CYP27B1遺伝子の応答性が、異なる転写制御因子をリクルートする複数のVDRE(ビタミンD応答エレメント)の異なる組み合わせによって、細胞タイプによって異なることが明らかにされました。
これらの研究は、ビタミンDの代謝と免疫系における役割を理解する上で重要な洞察を提供しており、特にビタミンDが自然免疫系に与える影響や、異なる細胞タイプでの遺伝子の発現制御の違いに関する理解を深めるのに貢献しています。
分子遺伝学
ビタミンD水酸化欠損型くる病、1A型
ビタミンD水酸化欠損型くる病、1A型(VDDR1A)は、1-α-ヒドロキシラーゼの活性不足により引き起こされる遺伝性疾患です。この酵素は、ビタミンDを活性化するために必要で、その活性不足はビタミンDの代謝異常、低カルシウム血症、くる病または骨軟化症の原因となります。
Fuら(1997年)の研究では、VDDR1A患者のケラチノサイトが1-α-ヒドロキシラーゼ活性を欠いていることが発見され、CYP27B1遺伝子のヌル変異の複合ヘテロ接合が特定されました。この発見は、VDDR1Aの分子基盤を理解する上での重要な進歩でした。
Kitanakaら(1998年)は、VDDR1A患者におけるホモ接合性ミスセンス変異を同定し、患者の家族内での遺伝的保因者の状態を明らかにしました。この研究は、疾患の遺伝的伝達に関する理解を深めました。
Wangら(1998年)の研究は、VDDR1A患者19人におけるCYP27B1遺伝子の多様な変異を同定し、民族間での変異の共有と独自のハプロタイプを発見しました。これは、VDDR1Aの分子遺伝学的多様性に光を当てるものです。
Kitanakaら(1999年)とWangら(2002年)の研究は、VDDR1Aのさらなる遺伝子変異を明らかにし、変異による機能的影響の範囲と、これが臨床的表現型に与える影響を解析しました。
Kimら(2007年)の研究は、10人の患者におけるCYP27B1遺伝子の新規変異と既知変異を同定し、VDDR1Aの遺伝子診断と臨床的診断に寄与しました。
これらの研究は、VDDR1Aの診断、治療、および遺伝相談に重要な情報を提供し、ビタミンD代謝疾患の理解を深める貴重な貢献をしています。
多発性硬化症との関連の可能性
Ramagopalanらによる2011年の研究では、多発性硬化症(MS)の発症者43人(MSに罹患している家系の4人以上)の全ゲノム配列が解析され、共通の機能喪失型バリアントや予測される損傷型バリアントは見つかりませんでした。しかし、CYP27B1遺伝子におけるヘテロ接合性の機能喪失型置換R389H(rs118204009)が1人の患者に見つかり、このバリアントは罹患家族4人全員(100%)および非罹患家族の33%に存在することが確認されました。このバリアントは、3,046人の親と罹患した子供の3人組(p = 1 x 10^(-5))およびさらに422組の親と罹患した兄弟姉妹(p = 0.046)の解析で過剰に伝播していることが示されました。R389H変異を持つ1人の患者の血清は、カルシトリオール濃度が低く、低いカルシトリオール濃度を持つMS患者96人のうち3人が推定される病原性CYP27B1バリアントを有していました。この結果は、MSリスクにおけるCYP27B1遺伝子の変異の原因的役割を支持します。
一方で、Banら(2013)およびBarizzoneら(2013)の研究では、CYP27B1遺伝子のR389HおよびL343FバリアントとMSとの間に有意な関連は見つかりませんでした。Banらは英国、米国、ノルウェーからの495の多重家族、2,092の単一罹患家族、および3,583人の対照群を含む4,594人のMS患者を比較し、Barizzoneらはイタリアとベルギーの患者2,608人と対照1,987人でR389HバリアントとMSとの関連を認めませんでした。両研究ともに、変異型CYP27B1対立遺伝子はMSの発症リスクに影響しないと結論付けています。
これらの研究結果は、CYP27B1遺伝子変異とMSの関連について異なる結論を示しており、この分野ではさらなる研究が必要であることを示唆しています。
動物モデル
Xueらによる2005年の研究では、Pth遺伝子やCyp27b1遺伝子を標的とした破壊を行ったマウスと、その両方の遺伝子が欠損している二重変異体を比較しました。Pth欠損マウスとCyp27b1欠損マウスは中程度の低カルシウム血症を示したのに対し、Pth/Cyp27b1二重欠損マウスは3週齢までに重度の低カルシウム血症で死亡しました。形態異常に関しては、Pth-nullマウスは最小限、Cyp27b1-nullマウスは骨端形成不全、Pth/Cyp27b1二重欠損マウスは重度の骨端形成不全を示しました。この研究からは、PTHと1,25(OH)2D3が骨格とカルシウムのホメオスタシスにおいて個別かつ協調的な役割を果たしていること、そして腎カルシウム輸送経路に協調的に影響を与えていることが示されました。
Vanhookeらによる2006年の研究では、Cyp27b1欠損マウスが1,25(OH)2D3を含むバランスの取れた食事で正常に成長することが確認されました。しかし、リンと1,25(OH)2D3が制限された環境では、これらのマウスは急速にくる病を発症しました。これは、ビタミンDとその代謝物の骨格への影響と、栄養が健康に与える影響の理解を深める研究です。
アレリックバリアント
.0001 ビタミンD水酸化欠損性くる病1a型
cyp27b1, arg107his
ビタミンD依存性くる病I型(264700)の日本人患者において、Kitanakaら(1998)は、CYP27B1遺伝子のエクソン2におけるホモ接合性のGからAへの転移を同定し、その結果、arg107からhis(R107H)への置換が生じた。In vitroでの機能発現研究により、変異型酵素は不活性であることが示された。
.0002 ビタミンd水酸化欠損性くる病1a型
cyp27b1, gly125glu
日本人ビタミンD依存性くる病I型(264700)患者において、Kitanakaら(1998)はCYP27B1遺伝子のエクソン2におけるホモ接合性のG-A転移を同定し、その結果、gly125-glu(G125E)置換が生じた。In vitroでの機能発現研究により、変異型酵素は不活性であることが示された。
.0003 ビタミンd水酸化欠損性くる病1a型
cyp27b1, arg335pro
ビタミンD依存性くる病I型(264700)の日本人患者において、Kitanakaら(1998)は、CYP27B1遺伝子のエクソン6にホモ接合性のGからCへの転座を同定し、その結果、arg335からpro(R335P)への置換が生じた。In vitroでの機能発現研究により、変異型酵素は不活性であることが示された。
.0004 ビタミンd水酸化欠損性くる病1a型
cyp27b1, pro382ser
ビタミンD依存性くる病I型(264700)の日本人患者において、Kitanakaら(1998)は、CYP27B1遺伝子のエクソン7にホモ接合性のC-T転移を同定し、pro382-to-ser(P382S)置換をもたらした。In vitroでの機能発現研究により、変異型酵素は不活性であることが示された。
.0005 ビタミンd水酸化欠損性くる病1a型
CYP27B1、1bp欠失、コドン211
ビタミンD依存性くる病I型(264700)の白人女児のケラチノサイトから得られたCYP27B1 cDNAにおいて、Fuら(1997)は2つの変異の複合ヘテロ接合を同定した:コドン211に1-bpの欠失があり、その結果、234位で早発停止コドンとなり、コドン231に1-bpの欠失があり、その結果、234位で早発停止コドンとなる(609506.0006)。これらの遺伝子によって産生されるタンパク質はひどく切断され、P450触媒活性を持たないであろう。
.0006 ビタミンd水酸化欠損性くる病1a型
cyp27b1、1-bp欠失、コドン231
Fuら(1997)によるビタミンD依存性くる病I型(264700)患者において複合ヘテロ接合状態で見つかったCYP27B1遺伝子のコドン231における1-bp欠失については、609506.0005を参照。
.0007 ビタミンD水酸化欠損性くる病1a型
cyp27b1, 1-bp 欠失, 958g
フランス系カナダ人のビタミンD依存性くる病I型(264700)患者10アレリック中9アレリックにおいて、Wangら(1998)はCYP27B1遺伝子の1-bp欠失(958delG)を同定した。ハプロタイプ解析は創始者効果と一致した。他の1例は米国の白人患者で、958delG変異のホモ接合体であった。
.0008 ビタミンd水酸化欠損性くる病1a型
cyp27b1, 7-bp dup
1α水酸化酵素の遺伝子は、438-442残基(pro-thr-pro-his-pro)をコードするエクソン8に5-prime-CCCACCC CCCACCC-3-primeの重複配列を持つ。Wangら(1998)は、ビタミンD依存性くる病I型(264700)の6家系の罹患者において、この7bpの重複配列を2つではなく3つ発見した。この変化はコドン455のヘムチオール酸システインの上流にあるため、得られた切断タンパク質は活性を持たない。家族は多様な民族(フィリピン人、ポーランド人、中国人、白人(米国)、黒人(米国)、ヒスパニック)で、変異の背景としていくつかの異なるハプロタイプが観察された。その結果、7bpの重複はいくつかの独立したde novoイベントによって生じたことが示唆され、ホットスポットであることが示された。
.0009 ビタミンd水酸化欠損性くる病1a型
cyp27b1, thr321arg
ビタミンD依存性くる病I型(264700)の軽度の臨床症状を有する患者において、Kitanakaら(1999)は、CYP27B1遺伝子の2つの変異の複合ヘテロ接合を同定した:エクソン5のCからGへの転位はthr321からargへの置換(T321R)をもたらし、イントロン3の最初のヌクレオチド(609506.0010)のGからAへの転位はイントロンの保持を引き起こすスプライシング変異をもたらした。In vitroでの機能発現研究では、どちらの変異型酵素も活性が残存しないことが示された。
.0010 ビタミンD水酸化欠損性くる病1a型
CYP27B1、IVS3DS、G-A、+1
北中ら(1999)によるビタミンD依存性くる病I型(264700)の患者において複合ヘテロ接合状態で発見されたCYP27B1遺伝子、609506.0009参照。
.0011 ビタミンD水酸化欠損性くる病1a型
cyp27b1, thr409ile
フィリピン人のビタミンD依存性くる病I型(264700)患者2人において、Wangら(1998)はCYP27B1遺伝子のエクソン8に3299C-T転移を同定し、thr409-to-ile(T409I)置換をもたらした。この変異は7bpの重複(609506.0008)との複合ヘテロ接合で生じた。
VDDR Iの患者において、Wangら(2002)はT409I変異をホモ接合で、またarg389からglyへの変異(R389G; 609506.0015)との複合ヘテロ接合で発見した。
.0012 ビタミンd水酸化欠損性くる病1a型
CYP27B1、ARG389HIS (rs118204009)
ビタミンD依存性くる病Ⅰ型(264700)の血縁関係のない2家族の罹患者において、Wangら(1998)はCYP27B1遺伝子の2つの変異、すなわちarg389からhisへの置換(R389H)と1bpの欠失の複合ヘテロ接合を同定した(609506.0007)。1家族は米国人で、もう1家族はフランス系カナダ人であった。
VDDR Iの患者において、Wangら(2002)はR389H変異を5-bp欠失/6-bp挿入(609506.0013)とleu343-to-phe変異(L343F; 609506.0016)との複合ヘテロ接合で発見した。R389H変異はP450c1-αタンパク質のβシート-1の4本鎖に起こり(Wang et al., 1998)、完全に不活性なタンパク質となる(Wang et al., 2002)。
多発性硬化症との関連の可能性
Ramagopalanら(2011)は、多発性硬化症(MS; 126200)の発端者43人(4人以上がMSに罹患している家系)の全ゲノム配列を決定し、1人の患者がCYP27B1遺伝子にヘテロ接合性のR389H置換(rs118204009)を有することを発見した。このバリアントは、3,046人の親と罹患した子供の3人組の解析(p = 1 x 10(-5))と、さらに422組の親と罹患した兄弟姉妹の解析(p = 0.046)でも過剰伝播していることが判明した。どの個体もVDDR1Aを認めなかった。フランス系カナダ人はいなかった。R389H変異を有する1人の血清は対照と比較してカルシトリオール濃度が低く、これは機能喪失対立遺伝子のヘテロ接合性がカルシトリオール濃度を低くすることを示唆している。全体として、この所見は、MSリスクにおけるCYP27B1遺伝子の変異の原因的役割を支持するものであり、疾患リスクに影響すると思われる地理的緯度勾配と相関していた。
Banら(2013年)は、495の多型家族、2,092の単一罹患家族、および3,583人の対照群と比較した4,594人のMS患者において、R389HバリアントとMSとの間に有意な関連を認めなかった。家族性の症例ではバリアントを有するものはいなかった。患者5人(0.07%)と対照2人(0.6%)がバリアントに対してヘテロ接合体であった。Barizzoneら(2013年)もまた、イタリアとベルギーの患者2,608人と対照1,987人において、R389HバリアントとMSとの関連を見いださなかった。ヘテロ接合のR389Hバリアントを有するMS患者1人と非罹患者1人の血漿測定では、1,25-ジヒドロキシビタミンD値の低下は認められなかった。Banら(2013)とBarizzoneら(2013)は、変異型CYP27B1対立遺伝子はMSの発症リスクに影響しないとそれぞれ結論づけた。
.0013 ビタミンD水酸化欠損性くる病、1a型
CYP27B1、5-bp欠失/6-bp欠失
Wangら(2002)は、ビタミンD依存性くる病(264700)の2人の患者において、CYP27B1遺伝子のエクソン2のヌクレオチド897から始まるGGGCGの欠失とCTTCGGの置換を発見した。この置換は読み枠のシフトを引き起こし、その結果コドン332で早発停止し、酵素活性を持たないペプチドとなった。1人の患者(米国の黒人少女)では、この変異はarg389からhisへの複合ヘテロ接合(R389H; 609506.0012)で起こり、もう1人の患者(ベルギーの少女)ではleu343からpheへの複合ヘテロ接合(L343F; 609506.0016)で起こった。
.0014 ビタミンd水酸化欠損性くる病1a型
cyp27b1, ivs2, g-a, +1
血縁関係のある両親から生まれたビタミンD依存性くる病(264700)のパキスタン人男児において、Wangら(2002)はイントロン2の最初のヌクレオチド(IVS2+1G-A)にホモ接合性のGからAへの転移を同定した。この変異は最初のドナー部位を破壊し、その結果イントロン2が保持されると著者らは予測した。ミニ遺伝子解析の結果、イントロン2の保持は読み枠のシフトと翻訳の早期終結を引き起こし、酵素活性のないペプチドをもたらした。
.0015 ビタミンD水酸化欠損性くる病1a型
cyp27b1, arg389gly
ビタミンD依存性くる病(264700)のチリ人女児において、Wangら(2002)はCYP27B1遺伝子のエクソン7にarg389からgly(R389G)に置換するCからGへの転座を発見した。この変異はT409I(609506.0011)との複合ヘテロ接合で生じた。R389G変異体タンパク質は1-α-ヒドロキシラーゼ活性を示さなかった。
.0016 ビタミンd水酸化欠損性くる病1a型
CYP27B1、LEU343PHE (rs118204011)
ビタミンD依存性くる病(264700)の軽症型であるベルギーの小児において、Wangら(2002)はCYP27B1遺伝子のエクソン6に2605C-T転移を同定し、leu343-to-phe(L343F)置換をもたらした。この変異はR389H(609506.0012)との複合ヘテロ接合で生じた。L343F変異は野生型酵素活性の2.3%を保持していた。
多発性硬化症との関連の可能性
Ramagopalanら(2011)は、多発性硬化症(MS;126200)の親-患児3人組のコホート3,046例においてL343F対立遺伝子の過剰伝達を発見し、このバリアントのヘテロ接合がカルシトリオール低値を引き起こしているという仮説を立てた。このバリアントは1873人の対照群では認められなかった。この所見は、MSリスクにおけるCYP27B1遺伝子の変異の原因的役割を支持するものであり、疾患リスクに影響すると思われる緯度勾配と相関していた。
Banら(2013)は、L232Fバリアント(rs118204011)とMSとの関連を、495の多重家族、2,092の単一罹患家族、および3,583人の対照群に対する4,594人の患者において見いださなかった。このバリアントを有する患者は1例のみであった。
.0017 ビタミンd水酸化欠損性くる病1a型
cyp27b1, glu189gly
ビタミンD依存性くる病(264700)の軽症型であるスイスの小児において、Wangら(2002)はCYP27B1遺伝子のエクソン3における1772A-G転移に起因するホモ接合性のglu189-to-gly(E189G)アミノ酸置換を同定した。この変異型タンパク質は野生型の22%の1-α-水酸化酵素活性を保持しており、in vitroで部分的な活性を保持した最初のミスセンス変異であると報告された。
多発性硬化症との関連の可能性
Ramagopalanら(2011)は、多発性硬化症(MS;126200)の親-罹患児3人組3,046人のコホートにおいてE189G対立遺伝子の過剰伝達を発見し、このバリアントのヘテロ接合性がカルシトリオールレベルの低下を引き起こしているという仮説を立てた。このバリアントは1873人の対照群では認められなかった。この所見は、MSリスクにおけるCYP27B1遺伝子の変異の原因的役割を支持するものであり、疾患リスクに影響すると思われる地理的緯度勾配と相関していた。
Banら(2013年)およびBarizzoneら(2013年)は、変異型CYP27B1対立遺伝子はMSの発症リスクに影響しないとそれぞれ結論している;609506.0012および609506.0016を参照のこと。







