目次

AHCY遺伝子がコードするアデノシルホモシステイナーゼ(SAHH)は、DNAやヒストンへのメチル基転移後に必ず生成される代謝副産物「SAH(S-アデノシルホモシステイン)」を分解できる、哺乳類唯一の酵素です。SAHが蓄積すると、100種類以上のメチル化反応がすべて停止します。そのため、AHCYは細胞のエピジェネティック状態を守る「代謝の守門者」と位置づけられており、その機能不全は発達障害・肝疾患・発がんリスクへと直結する広範な波及効果をもたらします。
Q. AHCY遺伝子とはどのような遺伝子ですか?まず結論だけ知りたいです
A. AHCY遺伝子は、1炭素代謝の要であるSAH(S-アデノシルホモシステイン)加水分解酵素(SAHH)をコードします。この酵素はDNA・RNA・ヒストンへのメチル基転移後に必ず生成されるSAHを除去できる哺乳類唯一の酵素であり、細胞のメチル化ポテンシャルを維持する絶対的な鍵を握っています。変異による機能不全は、SAHH欠損症(OMIM: 613752)という常染色体劣性の超稀少代謝疾患を引き起こします。
- ➤遺伝子の基本情報 → 染色体20q11.22・Gene ID 191・432アミノ酸・全身に普遍的に発現
- ➤1炭素代謝の中核 → SAH→アデノシン+ホモシステインへ分解、SAM/SAH比(メチル化ポテンシャル)を維持
- ➤エピジェネティクス → DNMT1・PRC2と直接相互作用、OCT4-AHCY-PRC2軸で幹細胞の多能性を維持
- ➤関連疾患 → SAHH欠損症(世界15〜19例の超稀少疾患)・脳梗塞リスク・成人発症肝細胞癌
- ➤がんとの関連 → 大腸・乳・膀胱がんでの過剰発現、H19 lncRNA回路、AHCY阻害剤の開発
1. AHCY遺伝子の基本情報:遺伝子座・発現・タンパク質概要
AHCY(Adenosylhomocysteinase)は、バクテリアから植物・酵母・線虫・昆虫を経て哺乳類に至るまで、地球上のほぼあらゆる生物に保存された代謝酵素です。ヒトゲノムでは染色体20q11.22に位置し、NCBI Gene IDは191です。別名としてSAHH(S-Adenosyl-L-Homocysteine Hydrolase)とも呼ばれます。
| 項目 | 詳細 |
|---|---|
| 公式シンボル(HGNC) | AHCY |
| その他の呼称 | SAHH、SAHH_HUMAN |
| 遺伝子座 | 染色体20q11.22 |
| Gene ID(NCBI) | 191 |
| タンパク質サイズ | 432アミノ酸残基/ホモ四量体 |
| 主な発現組織 | 腎臓(RPKM 29.2)、甲状腺(RPKM 22.9)をはじめ全身で普遍的に発現 |
| 主な細胞内局在 | 細胞質・核内(DNA複製期および転写活性時のクロマチン上) |
| 関連疾患(OMIM) | SAHH欠損症(OMIM: 613752)/常染色体劣性 |
AHCYは腎臓や甲状腺で特に高発現しますが、肝臓はAHCY酵素活性が脳の約10倍に達し、全身の1炭素代謝の最大のハブ臓器です。また、AHCYは単に細胞質に浮遊する代謝酵素ではありません。近年の研究により、DNA複製フォークや転写が活発なクロマチン領域へ動的にリクルートされ、その場でSAHを迅速に除去するという、時空間的に精密な機能が明らかになっています。
💡 用語解説:SAH(S-アデノシルホモシステイン)とは
SAH(S-アデノシルホモシステイン)は、細胞内でメチル基(CH₃)が転移されるたびに必ず生成される副産物です。DNAのシトシン塩基、ヒストンタンパク質、RNAのキャップ構造、脂質、クレアチン——これらへのメチル化反応はすべてSAM(S-アデノシルメチオニン)からメチル基を受け取り、その残骸としてSAHを放出します。SAHはメチル基を供与したSAMに構造が極めて似ているため、メチル化酵素の活性部位を競合的に占拠し、メチル化反応を強力にブロックします。AHCYはこのSAHを速やかに分解することで、メチル化反応が滞りなく継続できる環境を守ります。
2. タンパク質の立体構造と触媒メカニズム
AHCYタンパク質は、生体内で同一サブユニット4つが集まったホモ四量体(Homotetramer)を形成して機能します(代表的な結晶構造PDB ID: 9PDH)。高解像度の構造生物学解析から、各サブユニット(432アミノ酸残基)は機能的に分化した3つのドメインを持つことが明らかになっています。
💡 用語解説:ホモ四量体とは
「ホモ(homo)」は「同一の」、「四量体(tetramer)」は「4つのサブユニットが集合した構造体」を意味します。AHCYは同じタンパク質分子が4個集まって一つの機能ユニットを形成しています。1つのサブユニットだけでは酵素として機能できず、4個が正確に組み合わさって初めて活性を持ちます。遺伝子変異によってサブユニットの形が変わると、四量体の組み立て自体が崩れてしまうことがあります。
3つのドメイン構造と動的コンフォメーション変化
各サブユニットは以下の3つのドメインから構成されます。
- ① 基質結合ドメイン(アミノ酸残基1〜181および355〜385):SAHやそのアナログ(リバビリンなど)を認識して結合する最大のドメイン(212残基)。
- ② 補酵素結合ドメイン(アミノ酸残基197〜351):NAD+を結合する155残基のドメイン。哺乳類ではNAD+がサブユニットの深部に埋め込まれており、外部溶媒から保護されています。
- ③ C末端ドメイン(アミノ酸残基386〜432):ヘリックス・ループ・ヘリックス構造を持ち、隣接するサブユニットへと伸びることで四量体の安定性と完全性を担保します。
💡 用語解説:NAD+(ニコチンアミドアデニンジヌクレオチド)とは
NAD+は多くの酵素が触媒反応に利用する補酵素(コファクター)です。AHCYの場合、NAD+はタンパク質のコア深部に強固に結合しており、触媒反応の中間段階で一時的に還元型(NADH)に変換されます。NAD+なしではAHCYは機能できません。この補酵素との結合が非常に安定しているため、NAD+はAHCYと「一体」として機能します。
AHCYの触媒反応は高度に動的で、基質(SAH)の結合・放出に伴い大規模なコンフォメーション変化(立体構造変化)を起こします。基質が結合していない「オープン状態」では、基質結合ドメインと補酵素結合ドメインの間に大きな溝が開いています。SAHが結合するとヒンジ領域が約18度回転して溝が閉じる「クローズド状態」へ移行し、続いて二量体間で約14度の回転が生じて複雑な触媒反応が進行します。反応完了後、アデノシンが放出されると酵素は再びオープン状態へ戻ります。
変異が立体構造に及ぼす影響:酵素活性の劇的な低下
特定のアミノ酸変異がAHCYタンパク質の構造と活性に及ぼす影響は、機能解析実験によって詳細に解明されています。以下の表は野生型(正常)と疾患患者から見出された変異型の酵素活性を比較したものです。
| 酵素タイプ | 変異の特徴 | Km(μM) | 相対活性 |
|---|---|---|---|
| 野生型(WT) | 正常な四量体 | 15.09 | 100% |
| p.Asp86Glu | 負の電荷を保持 | 16.4 | 70.2% |
| p.Asp86Gly | 負の電荷の喪失・凝集体形成 | 測定不可 | 15.5% |
| p.Arg49Cys | 異常なジスルフィド結合形成 | 測定不可 | 6.7% |
特筆すべきは86番残基の負の電荷の重要性です。グルタミン酸(Glu)への置換は活性の70%を維持しますが、電荷を持たないグリシン(Gly)への置換では活性が15%まで低下します。これは、86番残基の負の電荷がSAH加水分解の直接の触媒機能には関与しないとされていたにもかかわらず、酵素の機能的構造維持に絶対的に不可欠であることを証明しています。
3. 1炭素代謝の中核を担う:AHCYの生化学的役割
AHCYが触媒する反応式は次の通りです:SAH(S-アデノシルホモシステイン)+ H₂O → アデノシン(Adenosine)+ L-ホモシステイン(L-Hcy)。この反応が1炭素代謝サイクル全体にとって「要」である理由は、SAHそのものの強力なメチル化阻害活性と、分解産物の重要な下流代謝にあります。
💡 用語解説:1炭素代謝(ワンカーボンメタボリズム)とは
「1炭素代謝」とは、1個の炭素原子(特にメチル基:-CH₃)を細胞内で転移・再利用するための代謝経路の総称です。DNAの複製・修復に必要なプリンやチミジンの合成、メチオニン・システイン・セリンなどのアミノ酸代謝、DNAやヒストンのメチル化によるエピジェネティクス制御——これらすべてが1炭素代謝によって支えられています。葉酸(ビタミンB9)やビタミンB12が1炭素代謝の重要な補助因子です。AHCYは、この広大な代謝ネットワーク全体の「詰まり」を防ぐ排水口のような役割を果たしています。
メチル化ポテンシャル(SAM/SAH比):細胞の「メチル化力」を示す指標
細胞がどれだけメチル化反応を行う余力があるかは、SAM/SAH比(AdoMet/AdoHcy比)、すなわち「メチル基供与体SAMの量÷阻害剤SAHの量」で評価されます。健常者ではこの比率は約4.10±2.11に保たれています。AHCYの機能が低下してSAHが蓄積すると、この比率が急激に低下し、全細胞でのメチル化ポテンシャルが喪失します。
AdoMet/AdoHcy比(メチル化ポテンシャル)の比較
0.52
1.48
4.10
出典:Molecular Genetics and Metabolism誌(肝移植によるメチル化ポテンシャルの回復データより)
AHCYの分解産物が担う下流代謝
AHCYがSAHを分解して生じるアデノシンとホモシステインは、それぞれ重要な代謝経路へと流れ込みます。
🔵 アデノシンの行方
プリン(核酸の構成要素)の再利用経路(サルベージ経路)に取り込まれ、AMP→ADP→ATPへと変換されます。下の画像はプリン・ヌクレオチド合成経路の全体像です。細胞が分裂・増殖するためのエネルギー通貨(ATP)および核酸原料の供給に直接貢献します。
🟢 ホモシステインの行方
メチオニンサイクルでSAMに再変換されるか、トランス硫化経路でシステイン→グルタチオン(主要な抗酸化物質)の合成に利用されます。DNA複製に伴う酸化ストレスへの防御においても、AHCYは重要な役割を果たしています。
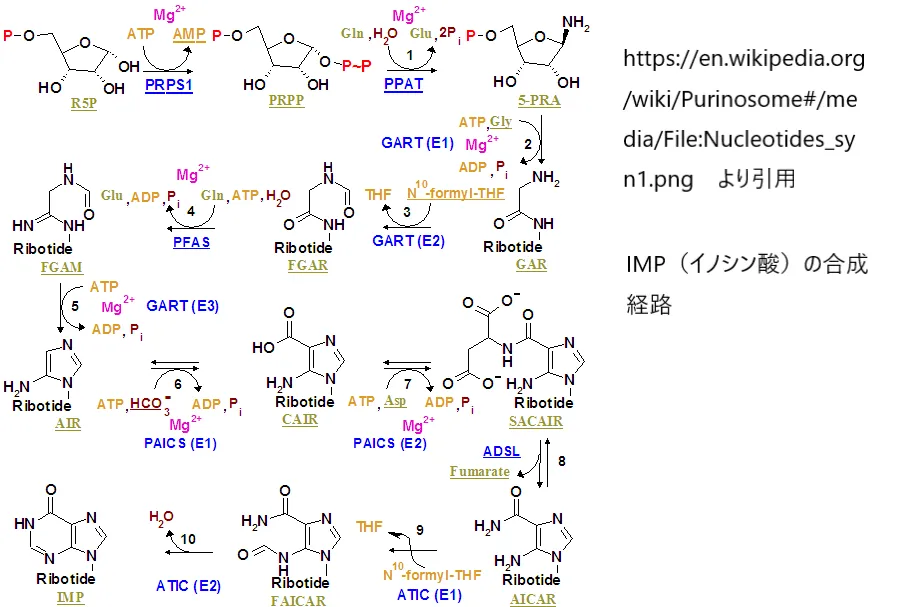
https://en.wikipedia.org/wiki/Purinosome#/media/File:Nucleotides_syn1.png
より引用
プリン・ヌクレオチド合成経路の全体像。AHCYが産生するアデノシンはこのサルベージ経路に組み込まれ、ATP・AMP等のプリンヌクレオチド供給に寄与します。
4. エピジェネティクス・幹細胞・発生における必須性
AHCYは単なる代謝酵素にとどまらず、細胞のエピジェネティック状態を時空間的に制御するマスターレギュレーターとしての機能を持ちます。近年の研究により、AHCYのクロマチンへの動的リクルートメントと複数のエピジェネティック機構との直接的な連携が明らかになっています。
💡 用語解説:エピジェネティクスとは
DNA塩基配列そのものを変えずに、遺伝子の「読まれ方(発現のオン・オフ)」を制御する仕組みの総称です。DNAのメチル化(特定の場所に-CH₃基を付加)やヒストン(DNAを巻きつけるタンパク質)の修飾が代表例です。エピジェネティクスの異常は、がん・発達障害・免疫疾患など多くの疾患と関連します。SAHはメチル化酵素を阻害するため、AHCYの機能不全はエピジェネティクスの全体的な崩壊を引き起こします。
DNMT1との直接複合体形成とDNA複製期のメチル化維持
細胞がDNAを複製するS期(合成期)において、AHCYはDNA維持メチル化酵素DNMT1と直接複合体を形成し、複製フォーク(DNAが実際に複製されている場所)に共局在します。in vitro(試験管内)の実験では、AHCYを加えるとDNMT1の触媒活性が直接増強されることが確認されています。逆に、薬剤やsiRNAでAHCYを阻害・ノックダウンすると、ゲノム全体の低メチル化(Hypomethylation)が誘導され、遺伝子発現パターンが根本から乱れます。
AHCYはそのC末端を介してmRNA(グアニン-N7)-メチルトランスフェラーゼ(CMT)とも直接結合します。CMTはmRNAの5’キャップ構造の最終メチル化を担う酵素であり、AHCYの阻害は未成熟なmRNAが核外へ異常に搬出される事態を引き起こします。AHCYはDNAメチル化だけでなく、RNAプロセシングと核外輸送にも深く関与しています。
OCT4-AHCY-PRC2軸:幹細胞の多能性を守る精密なバランス
マウス胚性幹細胞(mESC)において、幹細胞の自己複製マスターレギュレーターOCT4がAhcyプロモーターに直接結合してその発現を促進することが発見されました。この結合部位は非標準的モチーフ(ATTCAAAT)であり、デュアルルシフェラーゼアッセイでモチーフを変異させるとプロモーター活性が有意に低下します。
OCT4によって発現誘導されたAHCYは、幹細胞内で以下の2つの相反するエピジェネティック機能のバランスを精妙に維持します:
① 転写の活性化(H3K4me3)
大量のメチル化を必要とするH3K4me3(ヒストンH3の4番リジンのトリメチル化:活性化マーク)依存性遺伝子群の転写を、局所的なSAHの迅速除去を通じてサポートします。
② 転写の抑制(PRC2・H3K27me3)
ポリコーム抑制複合体2(PRC2)と直接相互作用し、中内胚葉分化遺伝子群へのH3K27me3(抑制マーク)によるサイレンシングを強固に維持します。
この「OCT4-AHCY-PRC2」軸により、幹細胞は自己複製能(多能性)を保ちながら不要な分化を防ぐという微妙なバランスを維持しています。AHCYの完全な欠失はマウスで胚性致死(着床前後の段階で発生が停止)をもたらし、この酵素が生命誕生の最初期から不可欠であることを示しています。

5. AHCY遺伝子変異と関連疾患
AHCY遺伝子の両アレル(2本の染色体の両方)に病的バリアントが生じると、「S-アデノシルホモシステインヒドロラーゼ欠損症(SAHH欠損症、OMIM: 613752)」という常染色体劣性遺伝の超稀少代謝疾患を引き起こします。
💡 用語解説:常染色体劣性遺伝とは
「常染色体」は性染色体(X・Y)以外の染色体のことです。「劣性(潜性)」とは、2本の染色体の両方に変異があって初めて発症する遺伝形式です。変異を1本だけ持つ人は「保因者(キャリア)」と呼ばれ、通常は無症状です。両親がともに保因者の場合、子どもが発症する確率は理論上25%です。SAHH欠損症はこの遺伝形式をとるため、両親がともに健康でも子どもが発症することがあります。
SAHH欠損症は世界で現在までに15〜19例程度しか報告されていない超稀少疾患(有病率:100万人に1人未満)です。疾患の根本原因はAHCY酵素の活性低下によるSAHの蓄積であり、その結果100種類以上のメチル化反応が広範に阻害されます。血漿中のSAM(AdoMet)は基準の1.7〜53倍(中央値21.5倍)、SAHは4.9〜193.8倍(中央値98.1倍)にまで上昇します。症状の詳細・治療法・遺伝カウンセリングについては、疾患専用ページをご覧ください。
中国の漢民族集団を対象とした研究では、AHCYプロモーター領域のDNAメチル化率上昇が虚血性脳卒中(脳梗塞)のリスクを有意に増加させることが示されています。AHCYプロモーターの高メチル化は酵素発現の低下を招き、ホモシステインの蓄積(高ホモシステイン血症)→血管内皮障害→血栓形成という連鎖を引き起こします。また、成人期(20〜30代)に初めてAHCY欠損症と診断された症例では、29歳で肝細胞癌を発症し32歳で死亡したという報告もあり、長期的なメチル化ポテンシャルの破綻が早期発がんを誘発する可能性が示唆されています。
6. がん生物学におけるAHCYの役割
AHCYの機能不全が稀少疾患を引き起こす一方で、がん細胞はしばしばAHCYを過剰に発現させ、自身の増殖・浸潤・生存に利用します。大腸がん・乳がん・胃のヘパトイド腺がん・膀胱尿路上皮がん(BLCA)など多くの腫瘍でAHCYの過剰発現が確認されており、腫瘍の悪性度・臨床病期の進行・全生存期間の短縮と強く相関しています。
p53状態に依存した細胞周期停止メカニズム
乳がん研究から、AHCYは発がん遺伝子c-MYCによって制御されることが示唆されています。siRNAを用いてAHCYをノックダウンすると、MEK/ERKシグナル経路のリン酸化が抑制され、強力な抗増殖効果と遊走・浸潤抑制効果が現れます。さらに、がん細胞内のがん抑制遺伝子p53の変異状態によって、AHCY阻害後の細胞周期停止パターンが全く異なります。
p53野生型がん細胞(MCF7等)
AHCY阻害→ATMキナーゼ活性化→p53リン酸化→Cyclin B1分解・CDC2リン酸化抑制→G2/M期停止
p53変異型がん細胞(MCF7-ADR等)
AHCY阻害→Cyclin D1・CDK6の発現抑制→G1期停止
H19 lncRNA回路:がん細胞によるエピジェネティックな「乗っ取り」
がん細胞は巧妙な機構でAHCYの機能を制御します。長鎖ノンコーディングRNA(lncRNA)の一種であるH19がAHCYタンパク質と直接結合してその加水分解活性を阻害することが発見されました。H19によるAHCY機能阻害は局所的なSAH濃度の急上昇を招き、DNAメチルトランスフェラーゼ(特にDNMT3B)の働きを強力にブロックします。その結果、特定のゲノム領域が低メチル化されて発現パターンが変化し、がん細胞の増殖・浸潤能力が増強します。
💡 用語解説:lncRNA(長鎖ノンコーディングRNA)とは
通常のRNAはDNAの情報をタンパク質に伝える「使者」ですが、lncRNAはタンパク質に翻訳されない200塩基以上のRNAの総称です。にもかかわらず、遺伝子発現制御・クロマチン構造調節・タンパク質との相互作用など多彩な機能を持ちます。H19は代表的なlncRNAの一つで、がんの発生・進行において重要な役割を持つことが近年明らかになっています。
AHCY阻害剤:新しいクラスの抗がん薬として
AHCYがんにおける中心的役割から、この酵素を特異的に阻害する化合物の開発が活発化しています。RapidFire高スループット質量分析(HTMS)システムを用いた約50万化合物のスクリーニングから、SAH結合部位に競合的に結合する複数の新規AHCY阻害剤が同定されています。これらは大腸がん細胞株(HCT116等)において細胞内SAHレベルを上昇させ、強力な増殖抑制効果を示します。AHCYの選択的阻害は「代謝的+エピジェネティック二重阻害」という新カテゴリーの抗がん療法として期待されています。
7. パラログ遺伝子AHCYL1(IRBIT)との機能的対比
ヒトゲノムには、AHCYと構造的に高い相同性を持つパラログ遺伝子としてAHCYL1(別名:IRBIT)(染色体1p13.3)およびAHCYL2が存在します。AHCYL1は脳(RPKM 81.1)や十二指腸(RPKM 50.1)で広く発現しますが、その機能はAHCYとは全く異なります。
| 機能的特徴 | AHCY(SAH加水分解酵素) | AHCYL1/IRBIT(SAHセンサー) |
|---|---|---|
| 酵素活性 | 完全なSAH加水分解活性 | 加水分解活性なし(疑似酵素) |
| 多量体形成 | ホモ四量体 | AHCYとヘテロ多量体形成→AHCY活性を抑制 |
| SAHへの応答 | SAHを基質として分解 | SAHを検知し構造変化(センサー機能) |
| 細胞内シグナル | DNMT1・CMTと結合、直接メチル化支援 | PIK3C3阻害→オートファジー抑制、IP3R阻害→Ca²⁺シグナル制御、Tauタンパク質相互作用 |
AHCYL1の最も注目すべき特性は、SAHが単なる代謝副産物ではなく、細胞の運命を決定する能動的なシグナル分子として機能することを証明した点です。細胞内のSAH濃度が上昇するとAHCYL1がSAHを検知して結合し、PIK3C3(ホスファチジルイノシトール3-キナーゼ)の活性を阻害します。その結果、MTORC1非依存的なメカニズムでマクロオートファジー(細胞の自食作用)が強力に抑制されます。また、AHCYL1はIP3受容体(IP3R)に結合して小胞体からミトコンドリアへのカルシウムイオン移行を制御し、アポトーシス誘導にも関与します。アルツハイマー病関連タンパク質Tauとの相互作用も確認されており、神経科学領域での新たな研究が進んでいます。
8. AHCY遺伝子に関連する遺伝子検査
AHCY遺伝子の変異を調べる遺伝子検査は、主に以下の目的で行われます。①SAHH欠損症が疑われる患者の確定診断、②保因者(キャリア)スクリーニング、③原因不明の代謝異常・高ホモシステイン血症の精査。ミネルバクリニックでは、1炭素代謝・ホモシステイン代謝に関連するNGS(次世代シーケンス)パネルを複数ご提供しています。
🧪 広域代謝疾患総合代謝NGSパネル先天性代謝異常症を広く網羅。原因不明の代謝疾患の精査に対応
🧬 ホモシステイン代謝メチルマロン酸尿症・ホモシスチン尿症NGSパネルホモシステイン代謝関連遺伝子を重点的に解析するパネル
👶 胎児水腫NIHF包括的NGSパネル重症AHCY欠損症では胎児水腫を生じることがあり、このパネルでAHCYも対象
保因者(キャリア)スクリーニング
SAHH欠損症は常染色体劣性遺伝のため、保因者(キャリア)の親は通常無症状です。妊娠前に自分がAHCY遺伝子の変異保因者かどうかを調べておくことで、子どもへの遺伝リスクを事前に把握できます。米国人類遺伝学会(ACMG)および米国産婦人科学会(ACOG)は、民族や家族歴にかかわらず、すべてのカップルへの拡張型保因者スクリーニングの提供を推奨しています。
男性向け男性向け拡張型保因者スクリーニングパートナーとともに保因者リスクを確認
妊娠前妊娠前遺伝子検査のご案内妊娠前からできる遺伝リスク把握の選択肢を解説
また、重症SAHH欠損症では胎児水腫(非免疫性胎児水腫:NIHF)を呈するケースがあります。胎児水腫の原因精査として行うNIHF遺伝子検査の対象にもAHCYは含まれます。検査の選択や解釈については、ACMG・ACOGの推奨内容も参考にしてください。
よくある質問(FAQ)
🏥 AHCY遺伝子・代謝疾患について専門医にご相談ください
SAHH欠損症・1炭素代謝異常・保因者スクリーニングに関するご相談は
臨床遺伝専門医が在籍するミネルバクリニックへ。
参考文献
- [1] Matos A, et al. Functional and Pathological Roles of AHCY. Front Cell Dev Biol. 2021;9:654344. [PMC8044520]
- [2] NCBI Gene. AHCY adenosylhomocysteinase [Homo sapiens]. Gene ID: 191. [NCBI Gene]
- [3] Liu Y, et al. AHCYL1 senses SAH to inhibit autophagy through interaction with PIK3C3 in an MTORC1-independent manner. Autophagy. 2022;18(2):397-413. [PMC8942424]
- [4] Zhang X, et al. AHCY: A metabolic gatekeeper at the interface of methylation, redox balance, and cellular stress response. Redox Biol. 2025. [PMC12955632]
- [5] Turner MA, et al. Structural insights into the reaction mechanism of S-adenosyl-L-homocysteine hydrolase. Nat Struct Mol Biol. 1998;5:369-376. [PMC4647836]
- [6] Buist NR, et al. S-Adenosylhomocysteine Hydrolase (AHCY) Deficiency: Two Novel Mutations with Lethal Outcome. PLoS Genet. 2006;2(4):e43. [PMC2876820]
- [7] Orphanet. S-adenosylhomocysteine hydrolase deficiency. ORPHA:88618. [Orphanet]
- [8] OMIM #613752. Hypermethioninemia with deficiency of S-adenosylhomocysteine hydrolase. Johns Hopkins University. [OMIM]
- [9] Stender S, et al. The biochemical profile and dietary management in S-adenosylhomocysteine hydrolase deficiency. Mol Genet Metab. 2022;137(1-2):88-96. [PMC9249945]
- [10] Esteve JM, et al. H19 lncRNA alters DNA methylation genome wide by regulating S-adenosylhomocysteine hydrolase. Nat Commun. 2016;7:10569. [PMC4703905]
- [11] Shi Y, et al. Inhibition of S-adenosylhomocysteine hydrolase decreases cell mobility and cell proliferation through cell cycle arrest. Cell Signal. 2015;27(11):2086-2096. [PMC4548325]
- [12] Wang Y, et al. The Role of AHCY Expression in Bladder Urothelial Carcinoma: A Bioinformatics and Experimental Analysis. Front Oncol. 2025. [PMC11937648]
- [13] Baric I, et al. Adult-onset liver disease and hepatocellular carcinoma in S-adenosylhomocysteine hydrolase deficiency. Mol Genet Metab. 2016;117(3):307-312. [PMC4733618]
- [14] Li YY, et al. The values of AHCY and CBS promoter methylation on the diagnosis of cerebral infarction in Chinese Han population. Medicine (Baltimore). 2020;99(44):e22893. [PMC7607831]






