目次
VDJ再構成
V(D)J組み換えは、T細胞やB細胞の成熟の初期に発生するリンパ球にのみ起こる体細胞組み換えのメカニズムである。その結果、B細胞とT細胞にそれぞれ見られる非常に多様な抗体/免疫グロブリンとT細胞受容体(TCR)のレパートリーが生み出される。
VDJのセグメントはいくつあるのか?
VDJセグメントはいくつもあり、その組み合わせによって様々なV領域を作ることができる。
ヒトκ(カッパ)型軽鎖の場合、機能的なVκ遺伝子セグメントは約40個、Jκ遺伝子セグメントは5個あり、組み合わせとしては40×5=200種類のVκ領域が存在することになる。
ヒトλ(ラムダ)型軽鎖の場合、機能的なVκ遺伝子セグメントは約30個、Jκ遺伝子セグメントは4個あり、組み合わせとしては30×4=120種類のVλ領域が存在することになる。
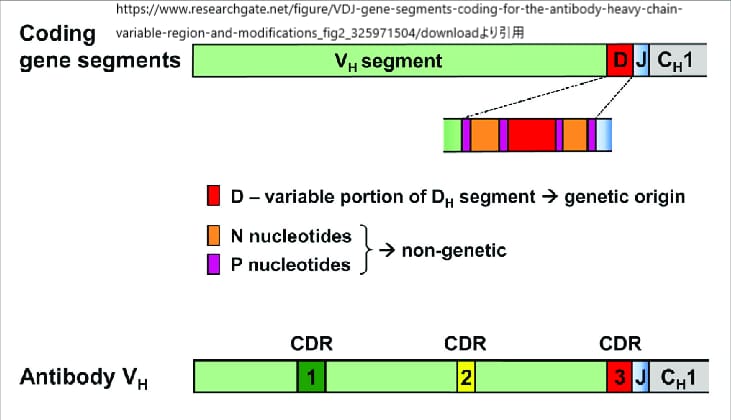
ヒトH鎖(重鎖)の場合、機能的なVH遺伝子セグメントは約40個、DH遺伝子セグメントが25個、JH遺伝子セグメントは6個あり、組み合わせとしては40×25×6種類のVκ領域が存在することになる。
V(D)J組換え
B細胞は多彩な化学構造に対する抗体を生成できる。この多様性がどのようにして生まれるのかについて謎がうまれた。この謎に対する答えは、意外にも非常に単純だった。
免疫グロブリンやT細胞受容体は2本のポリペプチド鎖(重鎖、軽鎖)からなり、双方が抗原結合部位を作っている。受容体の抗原結合部分をコードするエクソン(可変領域)は、発達中のリンパ球の染色体切断と再結合によって組み合わされる。抗原結合ドメインをコードするエキソンは、V(variable可変)、D(diversity多様)、J(joining接合)遺伝子セグメントから、DNAの切り貼りによる再配列の結果組み立てられている。一対のセグメントを選び、各セグメントに隣接して二本鎖切断を導入し、間にあるDNAを削除して、セグメント同士を結合させるこのプロセスは、V(D)J組み換えと呼ばれる。無数にある抗原に対応する多様性は、様々なセグメント間の接合部における少量のヌクレオチドの損失や獲得といった変化により、生来の持っている遺伝子のレパートリーが驚異的に増幅されることでもたらされ、潜在的には無限の抗原に特異的に結合できる抗体が生み出される。
Ig重鎖遺伝子、TCRベータまたはデルタ遺伝子の場合
Ig重鎖遺伝子、TCRベータまたはデルタ遺伝子の場合には完全な可変エキソンの組み立ては、2つのステップで行われる。
- 1. DとJのセグメントがいくつかの中から選ばれ、結合されてD-J再構成が形成される。
- 2. V領域が選ばれ、D-J再構成断片と結合して完全なVDJエクソンを形成する。
免疫グロブリン軽鎖遺伝子、TCRアルファおよびガンマ遺伝子の場合
免疫グロブリン軽鎖遺伝子、TCRアルファおよびガンマ遺伝子には、Dセグメントが存在しないので、V-J組み換えを伴う1ステップで再配列が行われる。
VDJ遺伝子組み換えの弱点
VDJ遺伝子再構成がなければ、無数の抗原に対する抗体をあらかじめ遺伝子として持っておく必要があり、新興感染症の病原体や想定外の毒物などを中和できないこととなり、生物は滅びてしまう。このため、VDJ再構成は非常に重要な役割を果たしている。
しかし、機能的な抗原受容体遺伝子を作り出すためには、染色体DNAを何度も切断しなければならない。つまり生物の一生で何百万回も切断しなければならないシステムであるがゆえに、大きなエラーを引き起こす可能性も内包している。V(D)J組み換えの異常は実際に起こり、この組換え異常が一般的なリンパ系新生物の発生を惹起することで、生命を脅かす可能性がある。リンパ系新生物のゲノム解析から、V(D)J組み換えによる発癌現象が明らかとなっている。
この記事の筆者:仲田洋美(医師)

- 総合内科専門医、臨床遺伝専門医、
がん薬物療法専門医
医療法人社団ミネルバ ミネルバクリニック 院長

この記事の筆者:仲田洋美(医師)






