目次
透明帯 zona pellucida

透明帯は、卵子の細胞膜の外側に存在する糖タンパク質の層である。卵子と精子の間の相互作用を制御し、多精子症を防ぐ。発育中の卵子を保護し、卵管への着床も防ぐ多彩な役割がある。
透明帯zona pellucidaとは?
透明帯zona pellucidaは、透明帯は、完全に成長した哺乳類卵母細胞の細胞膜を取り囲む厚い細胞の外側の被膜である。すべての哺乳類の卵と着床前胚を取り囲む細胞外被である。卵形成、受精、着床前の発育においてさまざまな重要な機能を果たしている。卵形成時には卵母細胞と卵胞細胞のコミュニケーションをサポートし、発育時には卵母細胞、卵、胚を保護し、受精時および受精後には排卵された卵と自由に泳ぐ精子との相互作用を制御している。透明帯を持たない卵を産む変異体のメスは不妊となる。現在、受精における透明帯の機能は、その糖タンパク質の一部に起因すると考えられている。
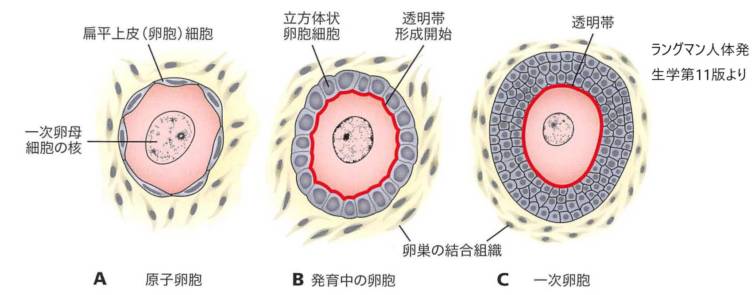
透明帯は、哺乳類の卵母細胞の細胞膜を囲む糖タンパク質の層である。卵母細胞の構成要素として重要な役割を果たしている。透明帯は一枚岩の初代卵母細胞に初めて現れる。卵母細胞と卵胞の両方から分泌される。透明帯は放射状の冠に囲まれている。冠は、卵巣から放出された卵をケアする細胞で構成されている。
この構造は、精子を結合し、先体反応を開始するのに必要である。哺乳類で最もよく知られているマウスでは、帯状の糖タンパク質であるZP3が精子の結合に関与し、精子の細胞膜上のタンパク質に付着している。そしてZP3は、精子が先体胞の内容物を放出する先体反応の誘発に関与する。他の生物種で何が起こっているのかを正確に把握するには、さらに多くの透明帯タンパクが同定されているため、より複雑になっている。
ヒトでは、受精後5日目に胚盤胞が透明帯のハッチングを行う。透明帯のハッチングとは、受精胚の成長過程で起こる現象であり、胚盤胞が子宮壁に着床するためには,この透明帯を取り除く必要があるため、透明帯は退化して分解し、下層の絨毛細胞に置き換わる。この透明帯は卵子の成長と受精に不可欠なものである。
透明帯の構成成分
マウス卵母細胞の各透明帯には透明帯の乾燥重量の80%以上、卵母細胞の全タンパク質の約17%に相当するタンパク質が含まれていることがわかっている。単離された透明帯の電気泳動分析ではZP1、ZP2、ZP3と名付けられた3つのタンパク質のみが存在していた。
見かけの分子量は200,000(ZP1)、120,000(ZP2)、83,000(ZP3)であり、糖タンパク質であることが確認された。ZP1、ZP2、ZP3は、それぞれ透明帯の全タンパク質の約36%、47%、17%を占めている。還元剤の存在下で透明帯を溶解させると、ZP1は見かけの分子量が13万の分子に変化し、非還元状態ではジスルフィド結合で安定化したオリゴマーとして存在することが示唆される。
透明帯を構成するZPタンパクの構造
ZPタンパク質は、4つの主要な特徴を持つ共通の構造を持っている。
- (1)分泌タンパク質であることを示すN末端の分泌シグナルペプチド(SP)
- (2)2種類のジスルフィド結合を持つ8個または10個の不変のシステイン残基を含む260個のアミノ酸からなる保存された配列であるZPドメイン
- (3)コンセンサス・フリン切断部位(CFCS)と呼ばれるプロタンパク質分解酵素ファミリーのメンバーに対する認識部位
- (4) 単一の膜貫通(TM)ドメインを含むC末端プロペプチド(CTP)
これらの要素は、ZPサブユニットの分泌と組み立てに重要な役割を果たしている。
ZPタンパクの生体内での分布
マウスの3つのZPタンパク質は、すべて透明帯ドメイン(ZPD)を持っている。
ZP1(アミノ酸271-540)、ZP2(アミノ酸364-628)、ZP3(アミノ酸45-302)の3つのZPタンパクは、ZPDタンパク質の原型であり、実質的にすべての脊椎動物の卵の透明帯ZP、ならびに脊椎動物および無脊椎動物の多種多様な組織および器官に存在している。
ZPDは6億年以上前に誕生し、クラゲ、ハエ、線虫からヒトまで、さまざまな生物で、受容体から機械的変換器まで多様な機能を持つ数百種類のZPDタンパク質が同定されている。
哺乳類では、脳、心臓、腸、腎臓、肝臓、鼻、卵巣、膵臓、脾臓、舌などに存在している。ZPDタンパク質は、多くの上皮細胞の膜表面に存在し、味覚や嗅覚などの感覚の機能に関与している。ZPDに加えて、上皮成長因子(EGF)、スカベンジャー受容体Cys-rich(SRCR)、フォンウィルブランド因子(vWF)、プラスミノーゲンアップルN-terminal(PAN)、補体c1r/c1s、Uegf、Bmp1(CUB)、ホエイ酸性タンパク質(WAP)などのドメインを持つものが多い。ZPDタンパク質には、ベータグリカン、キューティクリン、ダンピー、エンドグリン、グリコプロテイン-2、ヘンシン、メソグレイン、nompA、オイコシン、テクトリン、ウロモジュリン、ボメログランジンなどがある。ZPDタンパク質の前駆体には、ほとんどの場合、疎水性aaまたはグリコシルホスファチジルイノシトール(GPI)アンカーからなる膜貫通ドメイン(TMD)が1つ含まれており、これらは分泌タンパク質では失われている。ZPDタンパク質をコードする遺伝子の変異は、難聴、血管疾患、腎臓疾患、癌、そしておそらく不妊症を含む重篤な病的疾患を引き起こす可能性がある。
ZP欠損症と女性因子不妊症、臨床的意義
暗色の透明帯、楕円形、または不規則な形状の透明帯といった透明帯異形は、正常な卵子と比較して、体外受精後の着床率や妊娠率の低下と関連することが示されている(Sauerbrunら2015)。しかし、透明帯の異形性は、卵細胞質内精子注入法(ICSI)による妊娠率には影響しない。妊娠結果における透明帯/卵子の形態の予測可能性はまだ確立されていない。
透明帯ZPの形態とその不妊への影響に関するいくつかの研究に加えて、ヒトのZPタンパクをコードする遺伝子の変異が女性不妊の要因としてどのような役割を果たしているかを調べる研究もいくつか行われている。
体外受精で受精しなかった女性と、体外受精で受精に成功した女性、および不妊症であることが確認された女性のZp1、Zp2、Zp3、およびZp4遺伝子を分析したところ、Zp1およびZp3遺伝子の配列変異が1.5倍に増加していた。Zp3遺伝子の配列には2つの一塩基置換が存在し、その頻度は対照となる受胎可能なグループに比べて高いことがわかった。これらのうちの1つは、遺伝子の上流制御領域(element IIA)の保存されたモチーフにマッピングされており、ZP遺伝子の発現レベルの変化が、生殖能力に悪影響を及ぼすマトリックス形成の変化につながる可能性が示唆された。また、不妊クリニックに通院している不妊女性を対象とした別の独立した研究では、Zp2およびZp3をコードする遺伝子の配列変異が、ZPで観察される最も頻度の高い形態異常と関連している可能性が示されている。異常なZP1が卵母細胞の周りの透明帯の形成を妨げていることも示唆しされている。しかし、ヒトのZPタンパク質をコードする遺伝子の変異が、透明帯の形態異常や体外受精時の受精障害にどのような役割を果たすのかをさらに詳しく解明するためには、さらなる研究が必要である。
ご紹介
不妊症遺伝子検査
ミネルバクリニックでは不妊の原因が遺伝子にないかを検査する不妊症遺伝子検査をご提供しています。
お子さんに恵まれず悩んでいる女性に不妊治療方針の最適化をもたらせると期待されています。
この記事の著者:仲田洋美医師
医籍登録番号 第371210号
日本内科学会 総合内科専門医 第7900号
日本臨床腫瘍学会 がん薬物療法専門医 第1000001号
臨床遺伝専門医制度委員会認定 臨床遺伝専門医 第755号

この記事の筆者:仲田洋美(医師)






