プロモーターとは?
プロモーターは、DNAからRNAを合成する過程である転写の開始に関与する遺伝子の上流領域を指します。この領域には、基本転写因子が結合し、転写が開始されるための重要な役割を果たします[1][17]。プロモーターは、遺伝子の発現をコントロールするために必須な領域であり、ヒストンバリアントやヒストン翻訳後修飾が集積していることが特徴です[5]。
プロモーターの構造は、原核生物と真核生物で異なります。原核生物の場合、特有のDNAの塩基配列がs因子によって認識され、転写が開始されます[2]。一方、真核生物では、転写制御因子が遺伝子の上流のエンハンサー配列に結合することで転写が制御されます[2][3]。真核生物のプロモーターには、TATAボックスやTFIIB認識領域など、転写開始点を含むイニシエーターや転写開始点上流に存在する特定の配列があります[8]。
プロモーターの活性は、その配列に結合する転写因子や転写調節因子によって調節されます。これらの因子は、プロモーター領域内の特定の配列に結合することで、転写の活性化または抑制を行います[14]。プロモーターの解析により、セルラーゼやキシラナーゼの生産に関与する複数の転写調節因子が発見されており、プロモーター領域内におけるその結合配列が決定されています[14]。
プロモーターの研究は、遺伝子の発現制御機構の理解や遺伝子工学における遺伝子発現の制御など、生命科学の多くの分野で重要な役割を果たしています。
- 参考文献・出典
-
[1] www.setsurotech.com/glossary/promoter/
[2] www.sc.fukuoka-u.ac.jp/~bc1/Biochem/transcrp.htm
[3] bsw3.naist.jp/ko-kato/res-transcription.html
[4] www.riken.jp/press/2014/20140219_2/index.html
[5] www.yodosha.co.jp/jikkenigaku/keyword/3127.html
[6] www.tmd.ac.jp/artsci/biol/pdf/geneteng.pdf
[7] www.tmd.ac.jp/artsci/biol/textbook/geneteng.htm
[8] bsd.neuroinf.jp/wiki/%E3%83%97%E3%83%AD%E3%83%A2%E3%83%BC%E3%82%BF%E3%83%BC
[9] nsgene-lab.jp/expression/rna_synthesis/
[10] ruo.mbl.co.jp/bio/product/epigenetics/article/gene-expression.html
[11] www.try-it.jp/chapters-15090/sections-15091/lessons-15105/point-3/
[12] lib.ruralnet.or.jp/nrpd/
[13] www.tmd.ac.jp/artsci/biol/textintro/Chapt9.htm
[14] bio.nagaokaut.ac.jp/~Ogasawaralab/research/research004.html
[15] repository.dl.itc.u-tokyo.ac.jp/record/5598/files/K-03509.pdf
[16] www.wdb.com/kenq/dictionary/promoter
[17] ja.wikipedia.org/wiki/%E3%83%97%E3%83%AD%E3%83%A2%E3%83%BC%E3%82%BF%E3%83%BC
プロモーター領域は、遺伝子の転写を開始するために必要なDNA配列の一部で、転写因子と呼ばれるタンパク質が結合することでRNAポリメラーゼがその遺伝子の転写を開始するサイトです。プロモーターは遺伝子の直上流、すなわち転写開始部位のすぐ前に位置しています。
プロモーターは、単にRNAポリメラーゼの結合サイトとして機能するだけでなく、転写の調節エレメントが結合する場所でもあります。これにより、特定の細胞タイプ、発達段階、または外部の環境信号に応じて遺伝子がどのように転写されるかが決定されます。
● プロモーター領域の機能
1. 転写の開始: プロモーターはRNAポリメラーゼがDNAに結合し、RNAの合成を開始する場所を提供します。
2. 調節エレメントの結合: さまざまな調節タンパク質がプロモーターに結合することにより、遺伝子の活性が調節されます。これには活性化因子や抑制因子が含まれ、遺伝子の転写レベルを上げたり下げたりすることができます。
● プロモーター領域の拡張
近年の研究では、プロモーター領域が単一の配列に限定されず、より広範な領域にわたることが明らかになってきました。これは「拡張プロモーター」と呼ばれ、特に高等生物では遺伝子の転写を調節するための追加のエレメントが含まれることが多いです。この拡張領域にはエンハンサーやサイレンサーといった他の調節配列が含まれることもあり、これらは遺伝子の転写をより精密に制御するのに役立ちます。
● 現在の理解と課題
プロモーターの正確な定義と機能の理解は進化しています。科学者たちは、プロモーターがどのようにして遺伝子の転写を調節し、細胞の特性と遺伝子の発現パターンにどのように影響するかをさらに解明しようとしています。また、異なる生物や条件でのプロモーターの挙動を理解することは、遺伝子治療やバイオテクノロジーなどの応用にも重要です。
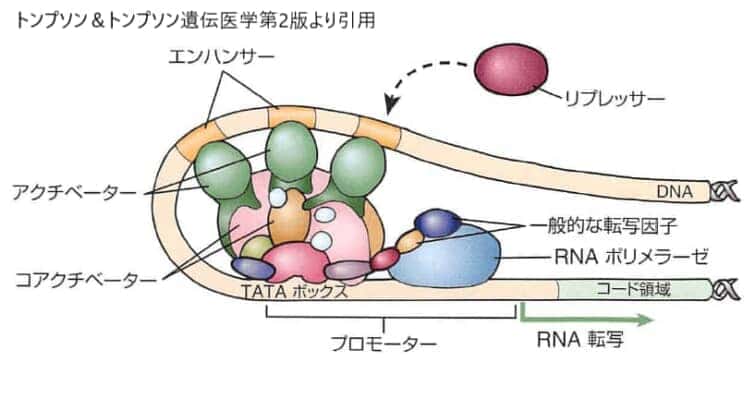
この図は一般的(基本的)な転写因子とRNAポリメラーゼをイメージにしたものです。転写因子とRNAポリメラーゼはメッセンジャーRNA(mRNA)の転写開始点に近接するシス作用性配列に結合します。ようなシス作用性配列をまとめてプロモーターと呼びます。これより遠位(3’側)のエンハンサーあるいはサイレンサーエレメントにはその配列に特異的な転写因子や組織特異的転写因子が結合します。コアクチベータータンパクは特異的転写因子と一般的な転写因子との間の生化学的相互作用を促進する働きがあります。
コアプロモーターとは?
コアプロモーターは、転写開始点の周辺、具体的には-50から+50ベースペア(bp)の領域に存在し、基本転写因子によって認識されることで、転写が生じる位置や向きを決定するDNAの領域です[1]。この領域は、RNAポリメラーゼIIによるmRNAの転写を担う遺伝子の正しい位置からの適切な転写開始に必要なコアプロモーターと、そのすぐ上流に存在してその本質的な活性を高める領域とがあります[3]。コアプロモーターには、TATAボックス、B認識配列(BRE)、イニシエーター(Inr)、下流コアプロモーター配列(DPE)などの主要なコンセンサス配列が含まれており、これらは基本転写因子が結合する配列です[3]。
コアプロモーターは、エンハンサーからの転写活性化の合図を受け取り、遺伝子発現を開始する役割を担っています。エンハンサーは、転写活性のON/OFFを切り替えるスイッチとして機能し、個体発生における遺伝子発現を時空間的に制御します。一方で、コアプロモーター自身も転写バーストの制御において重要な役割を果たすことが示されています[5]。
また、植物においては、哺乳類で知られている非TATA型プロモーターとしてのCpGアイランドを持つタイプは存在せず、代わりにGA型がそのニッチを埋めていることが見いだされています。これは、コアプロモーターのタイプが遺伝子機能や構造と関連しており、その違いが発現上の違いにとどまらず、遺伝子構造などにも影響を与えていることを示唆しています[1]。
- 参考文献・出典
-
[1] www.jstage.jst.go.jp/article/jspp/2009/0/2009_0_0065/_article/-char/ja/
[2] bsw3.naist.jp/ko-kato/res-transcription.html
[3] nsgene-lab.jp/expression/euk-polii_promoter/
[4] www.toho-u.ac.jp/sci/bio/column/0804.html
[5] www.u-tokyo.ac.jp/focus/ja/press/z0207_00054.html
[6] photosyn.jp/pwiki/
[7] ja.wikipedia.org/wiki/%E3%83%97%E3%83%AD%E3%83%A2%E3%83%BC%E3%82%BF%E3%83%BC
転写基本因子とは
転写基本因子(General transcription factor、GTF)とは、真核生物のクラスII遺伝子を鋳型mRNAへ転写する際に必要なタンパク質群のことです[7]。これらの因子は、RNAポリメラーゼIIがDNAのプロモーター領域を正確に認識し、転写を開始するために不可欠です。転写基本因子は、RNAポリメラーゼIIと共にコアプロモーター上に転写開始複合体を形成し、転写開始点からのRNA合成を促進します[5]。
真核生物のRNAポリメラーゼIIによる転写には、TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIHの6種類の基本転写因子が関与しています[4]。これらの因子は、以下のような役割を果たします:
– TFIID: コアプロモーター配列中に存在するTATAボックス、Inr、DPE、DCE、MTEなどを認識し、RNAポリメラーゼIIの転写開始に必要な構造を形成します。TFIIDはTATA結合タンパク質(TBP)とTBP関連因子(TAF)から構成されます[4]。
– TFIIA: TFIIDのTBPと相互作用し、TATAボックスへの結合を安定化させる役割を持ちます。
– TFIIB: TFIIDと相互作用し、RNAポリメラーゼIIの転写開始点への結合を促進します。
– TFIIE: RNAポリメラーゼIIの活性を調節し、TFIIHのヘリカーゼ活性を調節することで転写開始に必要なDNAの開裂を促進します。
– TFIIF: RNAポリメラーゼIIがプロモーターに結合する際の安定化を助け、転写開始後のポリメラーゼの進行を促進します。
– TFIIH: ヘリカーゼ活性を持ち、DNAの開裂を行い、RNAポリメラーゼIIのC末端ドメイン(CTD)のリン酸化を行うことで転写開始を促進します。
これらの基本転写因子は、転写調節因子とも相互作用し、転写の活性化や抑制を行うことで、細胞内での遺伝子発現の精密な調節を可能にしています[8]。転写基本因子の異常は、転写障害を引き起こし、様々な疾患の原因となることが知られています[1]。
- 参考文献・出典
-
[1] www.jstage.jst.go.jp/article/jscc1971b/27/2/27_52/_pdf/-char/ja
[2] www.sc.fukuoka-u.ac.jp/~bc1/Biochem/transcrp.htm
[3] www.saitama-med.ac.jp/jsms/vol28/04/t85_95.pdf
[4] nsgene-lab.jp/expression/pol2-gtf/
[5] bsd.neuroinf.jp/wiki/%E3%83%97%E3%83%AD%E3%83%A2%E3%83%BC%E3%82%BF%E3%83%BC
[6] www.try-it.jp/chapters-15090/sections-15140/lessons-15157/
[7] ja.wikipedia.org/wiki/%E5%9F%BA%E6%9C%AC%E8%BB%A2%E5%86%99%E5%9B%A0%E5%AD%90
[8] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-05273203/
プロモーターの種類:TATAプロモーターとTATAレスプロモーター
TATAボックスは、真核生物及び古細菌の遺伝子において、RNAポリメラーゼIIによる転写開始位置の上流25塩基対(bp)の位置、あるいはさらに上流に存在する共通した塩基配列で、通常は「TATAAA」の配列を持ちます[3]。この配列は、転写基本因子の一つであるTFIIDのサブユニットであるTATA結合タンパク質(TBP)によって認識され、転写開始部位を明らかにする目印として機能します[2]。TATAボックスは、特にタンパク質をコードする遺伝子のプロモーターに存在し、転写の効率化に寄与しています。
一方、TATAレスプロモーターは、TATAボックスを持たないプロモーターのことで、ハウスキーピング遺伝子などに多く見られます[3]。これらのプロモーターは、TATAボックスの代わりにGCボックスや下流プロモーターエレメント(DPE)などの他のコアプロモーター要素を持ち、複数の転写開始点を持つことも多いです[1]。TATAレスプロモーターは、細胞の基本的な生命活動に関わる遺伝子の発現を安定的に維持するために重要な役割を果たしています。
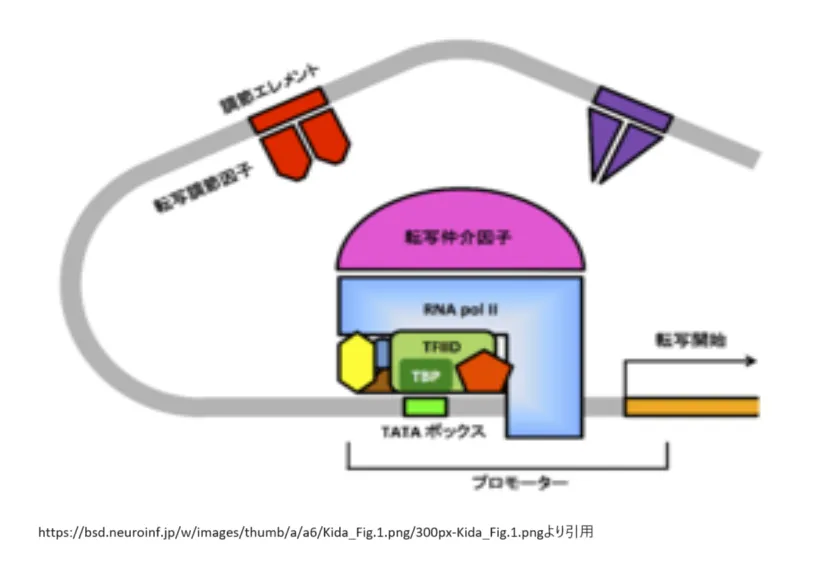
調節エレメントとは?
調節エレメント(regulatory elements)は、DNA上に存在する特定の配列であり、遺伝子の発現を調節する役割を持っています。これらのエレメントは、転写因子と呼ばれるタンパク質が結合することによって、RNAポリメラーゼの活性を促進または抑制し、遺伝子の転写をコントロールします[2][3][5][7][8]。
調節エレメントには、プロモーター、エンハンサー、サイレンサーなどが含まれ、それぞれが遺伝子発現の異なる側面を制御します。プロモーターは、転写開始点の近くに位置し、RNAポリメラーゼと転写基本因子が結合する場所です。エンハンサーは、プロモーターから離れた位置に存在することもあり、転写効率を高める役割を持ちます。サイレンサーは、遺伝子発現を抑制する機能を持つエレメントです[2][3][5]。
これらの調節エレメントは、細胞特異的な遺伝子発現や、環境や発達段階に応じた遺伝子発現の変化を可能にするため、生物の多様な生命現象を支える基盤となっています。また、調節エレメントは、遺伝子治療や分子生物学的研究においても重要な役割を果たし、遺伝子の発現パターンを理解するための鍵となる要素です[3][5][7].
- 参考文献・出典
-
[2] www.pha.fukuoka-u.ac.jp/seika
[3] wwwcrl.shiga-med.ac.jp/home/seminar/toku_sem/sp99/idenshi/home.html
[4] bsd.neuroinf.jp/wiki/%E8%BB%A2%E5%86%99%E5%88%B6%E5%BE%A1%E5%9B%A0%E5%AD%90
[5] bsd.neuroinf.jp/wiki/%E3%83%97%E3%83%AD%E3%83%A2%E3%83%BC%E3%82%BF%E3%83%BC
[6] www.toho-u.ac.jp/sci/biomol/glossary/bio/transcription_factor.html
[7] minerva-clinic.or.jp/academic/terminololgyofmedicalgenetics/tagyou/regulatory-elements/
[8] ja.wikipedia.org/wiki/%E3%82%B7%E3%82%B9%E3%82%A8%E3%83%AC%E3%83%A1%E3%83%B3%E3%83%88
遺伝子の転写調節機構とは?
転写調節機構とは、細胞内で遺伝子の転写がいつ、どの程度行われるかを制御する一連のプロセスです。この機構は、遺伝子の発現を組織特異的、発生段階特異的、または環境条件に応じて調節することで、細胞の機能と生物の発達をコントロールします[1]。
転写調節機構には、以下の要素が含まれます:
– プロモーター: RNAポリメラーゼIIが結合し、転写を開始するDNA上の領域で、基本転写因子が結合して転写開始を調節するコアプロモーター、組織や細胞ごとに異なる転写因子が結合する近傍プロモーターなどがあります[5][6]。
– 転写因子: 特異的DNA配列に結合し、RNAポリメラーゼによる転写を促進または抑制するタンパク質群です。これらは、エンハンサーやサイレンサーなどの転写制御配列に結合し、転写の正負を制御します[4]。
– エンハンサーとサイレンサー: 転写を活性化(エンハンサー)または抑制(サイレンサー)するDNA配列で、転写制御因子が結合する場所です。
– エピジェネティックな制御: DNAメチル化、ヒストン修飾などの化学的変更により、遺伝子のアクセシビリティと転写活性を調節するプロセスです。
転写調節機構は、細胞の応答性と適応性を高めるために、複雑で精密な制御が行われるシステムです。例えば、細胞がストレスを受けたときや、発達の特定の段階にあるときに、必要な遺伝子が適切なタイミングでオンまたはオフになるように調節されます。また、代謝産物がRNAに結合してその動態を制御するリボスイッチのようなメカニズムも存在することが示唆されています[3]。
- 参考文献・出典
-
[1] www.jstage.jst.go.jp/article/nogeikagaku1924/64/4/64_4_918/_pdf/-char/ja
[2] altair.sci.hokudai.ac.jp/g6/Research/TRCFs.html
[3] seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2016.880308/data/index.html
[4] bsd.neuroinf.jp/wiki/%E8%BB%A2%E5%86%99%E5%88%B6%E5%BE%A1%E5%9B%A0%E5%AD%90
[5] www.toho-u.ac.jp/sci/bio/column/0804.html
[6] bsd.neuroinf.jp/wiki/%E3%83%97%E3%83%AD%E3%83%A2%E3%83%BC%E3%82%BF%E3%83%BC
[7] nsgene-lab.jp/expression/bac_regulation2/
[8] www.jstage.jst.go.jp/article/jcrsj1959/45/1/45_1_19/_pdf







