目次
エンドフィリンは細胞のエンドサイトーシス過程において重要な役割を果たすタンパク質です。この記事では、エンドフィリンの機能、特に神経細胞におけるその役割とシナプス機能への影響について詳しく解説します。
第1章:エンドフィリンとは
エンドフィリンの基本的な特性
エンドフィリンは、細胞内膜動態に関与する進化的に保存されたタンパク質ファミリーで、5つのアイソフォームが存在し、様々な組織に分布しています。このタンパク質は、小胞エンドサイトーシス、ミトコンドリアの形態変化、アポトーシス、オートファゴソーム形成など、多岐にわたる生物学的機能を担っています。最近の研究では、エンドフィリンが神経変性疾患、腫瘍、心血管疾患、自己免疫疾患などの疾患の発症と進行に関与していることが示されており、疾患治療への応用が期待されています。
エンドフィリンは、細胞内輸送や細胞膜の再構成に関わるタンパク質で、特にエンドサイトーシスに重要な役割を果たします。細胞膜から小胞を形成し細胞内に取り込む過程で、膜の曲げや小胞の切り離しを促進します。また、アクチン結合タンパク質として細胞骨格と相互作用し、シナプス機能の調節やシグナル伝達の調節にも関与します。さらに、エンドフィリンの機能不全や異常は、神経系の発達障害や神経退行性疾患などに関連する可能性があります。
エンドフィリンの発見と研究の歴史
エンドフィリンの発見は、シナプス小胞が神経伝達物質の放出に重要であることを示す研究によって支えられています。1960年代、Bernard Katzらによる神経−筋接合部での電気生理学的実験と、Paladeらによる電子顕微鏡での脳組織観察がシナプス小胞の存在を明らかにしました。1980年代には、HeuserとReeseらがシナプス小胞と形質膜の融合像を電子顕微鏡下で観察し、エキソサイトーシスによる神経伝達物質の放出メカニズムを提唱しました。
エンドフィリンの構造
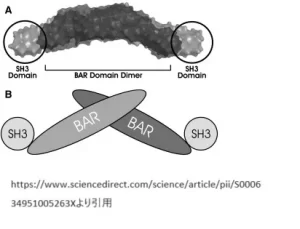
エンドフィリンファミリーには、特別な構造を持つタンパク質が含まれています。これらはSH3という部分とBARという部分から構成されていて、1996年に初めて見つかりました。エンドフィリンAというタイプには、A1、A2、A3の3種類があります。一方、エンドフィリンBには、B1とB2の2種類があります。
https://www.sciencedirect.com/science/article/pii/S000634951005263X
より引用
●SH3とは「Src Homology 3」という名前のタンパク質ドメインです。このドメインは約60アミノ酸から構成されており、特定のプロリン豊富な配列と結合する能力を持ちます。SH3ドメインを持つタンパク質は、細胞のシグナル伝達、細胞骨格の組織、エンドサイトーシス(細胞が外部の物質を取り込むプロセス)など、多くの生物学的プロセスに関与しています。
SH3ドメインの発見は、細胞内のシグナル伝達経路の理解に大きな貢献をしました。これは、多くのタンパク質間相互作用が特定の短いアミノ酸配列を介して行われることを示しており、その中でもSH3ドメインはプロリン豊富な配列と結合することで、特定のタンパク質同士を引き合わせる役割を果たしています。
結合のメカニズム: SH3ドメインはプロリン豊富な配列を持つタンパク質、例えばシナプスタンパク質やダイナミンといったタンパク質と結合します。これらの結合は、細胞のシグナル伝達や物質輸送などのプロセスにおいて中心的な役割を果たします。
BARドメインとの連携: SH3ドメインは可変長のスプライス領域を介してBARドメインと結合します。この結合により、エンドフィリンが細胞膜の特定の領域に正確に局在することを助け、ターゲットタンパク質との相互作用を促進します。
●BAR(Bin/amphiphysin/Rvs)ドメインは、タンパク質の一部分であり、細胞膜の曲がりや変形を感知したり、影響を与えたりする機能を持っています。このドメインは特に、細胞膜におけるさまざまなプロセスに関与しているタンパク質に見られます。そのプロセスには、エンドサイトーシス(細胞が膜をくぼませて外部から物質を取り込む現象)、膜の再配置や細胞骨格の組織などが含まれます。
BARドメインを持つタンパク質は、その形状が特徴的で、膜を曲げる能力を持っていることが知られています。このドメインは、膜と相互作用することで、膜を特定の方向に曲げることができ、それによって細胞内での物質の動きや膜の形状の変化を促します。例えば、エンドサイトーシスの過程では、BARドメインを持つタンパク質が細胞膜を特定の形に曲げ、外部からの物質を細胞内に取り込むための小胞を形成するのに寄与しています。
BARドメインは、細胞のダイナミックな膜構造の形成と再構成に重要な役割を果たし、細胞の機能と応答に不可欠な要素です。細胞のさまざまな生物学的プロセスを理解する上で、BARドメインとその機能は非常に重要な研究対象となっています。
N-BARドメイン
膜との結合: N-BARドメインは脂質二重層と直接結合し、細胞膜の湾曲を誘導する能力を持ちます。この特性は、細胞膜の動的な構造変化、特にエンドサイトーシスやシナプス小胞のリサイクリングにおいて重要です。
構造: N-BARドメインには、N末端の両親媒性ヘリックス(H0)、ヘリックス1(H1)に挿入された別の両親媒性ヘリックス、そして二量体化したBARドメインによって形成される三日月形のボディが含まれます。
H0: 膜結合を仲介します。
H1: 膜に挿入し、膜の湾曲を促進する役割を果たします。
二量体化と膜の湾曲: N-BARドメインは二量体化することにより、より強固な膜の湾曲を誘導し、安定化させます。このプロセスは、膜の形状変化を促進し、細胞内での多様な機能を可能にします。
エンドフィリンAは、細胞内で物質を取り込む役割を持つ過程を調整することが分かっています。特に、B1タイプは細胞の自滅プロセスに関わり、オートファジーという細胞が自らを清掃する過程も促進します。
エンドフィリンのサブタイプ
エンドフィリンA1、エンドフィリンA2、エンドフィリンA3、およびエンドフィリンB1とB2は、エンドフィリンファミリーの異なるメンバーであり、それぞれが異なる組織や臓器で特有の発現パターンを持ち、細胞内での特定の局在性を示します。
– エンドフィリンA1(別名SH3P4またはSH3GL2)は、主に脳組織で発現しており、神経系の機能に重要な役割を果たしていることが示唆されます。この発現パターンは、エンドフィリンA1が神経細胞の活動、特にシナプスの機能や神経伝達物質のリサイクリングに関わっていることを示しています。
– エンドフィリンA2(別名SH3P8、SH3GL1、EEN)は、膵臓、胎盤、前立腺、睾丸、子宮など多岐にわたる組織や臓器に発現しています。この広範な発現は、エンドフィリンA2が様々な生物学的プロセスや細胞タイプにおいて重要な役割を果たしていることを示しています。
– エンドフィリンA3(別名SH3P13またはSH3GL3)は、脳と精巣組織に主に発現しており、これらの組織特有の機能に関与している可能性があります。エンドフィリンA3の特定の役割については、さらに詳細な研究が必要ですが、神経系および生殖系におけるその重要性が示唆されています。
– エンドフィリンB1は、心臓、骨格筋、腎臓、胎盤で特に高い発現を示しています。細胞内膜、ミトコンドリア膜、ゴルジ膜、オートファゴソーム膜に局在し、これらの組織のエネルギー代謝、筋肉の動作、細胞の生存と死、およびオートファジーといったプロセスに重要な役割を担っています。
– エンドフィリンB2は、骨格筋、脂肪組織、肺、脳、乳腺に発現しています。この発現パターンは、エンドフィリンB2がこれらの組織における多様な生物学的機能に関与していることを示しています。
エンドフィリンファミリーのこれらのメンバーは、細胞内で異なる局在と発現パターンを持ち、細胞のシグナル伝達、物質輸送、代謝プロセスなどにおいて特異的な役割を果たしています。これらのタンパク質が正常に機能することで、細胞や組織の正常な生理を維持しています。
エンドフィリンAの機能
エンドフィリンAには、SH3とN-BARという二つの重要な部分があります。SH3は、小胞が細胞内に運ばれる際に他のタンパク質と結合するのに役立ちます。一方、N-BARは小胞の形状を変えるのを助けることで、物質の取り込みをスムーズにします。
最近の研究では、エンドフィリンAが細胞のオートファジーという過程にも関与している可能性が示されています。オートファジーは、細胞が不要な部分を分解して再利用することで、細胞の健康を保つ重要な機能です。エンドフィリンAは、この過程を促進することで、細胞がストレスに対応しやすくなることが期待されています。
さらに、エンドフィリンAは細胞がどのようにして特定の物質を迅速に取り込むか、つまり「高速エンドフィリン媒介エンドサイトーシス(FEME)」にも重要です。この過程は、細胞が必要とする物質を素早く取り込むためのもので、エンドフィリンAはこの過程で中心的な役割を果たします。
簡単に言うと、エンドフィリンAは細胞が生きるために必要な物質を取り込んだり、細胞内で物質を運んだりするのに重要なタンパク質であり、細胞の健康維持に欠かせない存在です。
エンドフィリンBの機能
エンドフィリンBについても分かりやすく説明しましょう。
エンドフィリンB1(またはBif-1)は、細胞の自然な死であるアポトーシス、ミトコンドリアの形状変更、そしてオートファゴソーム(細胞内の不要物を分解するための構造)の形成に関わる重要なタンパク質です。最初にアポトーシスを促進するタンパク質として発見されたBif-1は、特にミトコンドリアでアポトーシスを調節する新しい役割を持っていることが分かっています。
アポトーシスが起こると、Bif-1は細胞質からミトコンドリアへ移動し、そこでオリゴマー(複数のタンパク質が結合した構造)を形成します。このオリゴマーは細胞膜を変形させ、アポトーシスを進めるためのリポソーム小胞を形成します。Bif-1はまた、Baxというタンパク質と結合し、これがミトコンドリアに移動してアポトーシス関連因子、例えばチトクロームCの放出を促進します。
Bif-1の別の役割はオートファジーの誘導です。オートファジーは細胞が自らの一部を分解し、栄養を再利用する過程であり、細胞の健康維持に不可欠です。Bif-1は、ベクリン-1という別のタンパク質と複合体を形成し、これによりオートファジーが促進されます。この過程では、細胞内の動的な膜構造が再配列され、オートファゴソームが形成されます。これにより、細胞は不要なタンパク質や小器官を効率的に分解して、飢餓時などに必要な栄養素を供給することができます。
Bif-1はオートファジーの初期段階、特にオートファゴソームの膜が形成される過程に関わっています。また、Bif-1はオートファジーの標識となるタンパク質、Atg5やLC3とも共局在することが確認されており、オートファゴソーム形成におけるその役割をさらに強調しています。
要するに、エンドフィリンB1は細胞の自然死であるアポトーシスを促進し、ミトコンドリアの形状を調節し、オートファジーを通じて細胞の健康を維持する多機能のタンパク質です。これらの過程はすべて、細胞が正常に機能し、生存するために重要です。
第2章:エンドフィリンの機能とメカニズム
エンドサイトーシスにおけるエンドフィリンの役割
エンドサイトーシスは細胞が外部から物質を取り込むプロセスであり、細胞膜が内側にくぼみ、取り込みたい物質を包み込む小胞を形成することで行われます。エンドフィリンはこのプロセスにおいて重要な役割を担っています。
エンドフィリンは、特にSH3ドメインとBARドメインを持つタンパク質で、この2つのドメインがエンドサイトーシスにおけるその機能を果たすために重要です。
1. BARドメインは膜の曲げに関与しており、エンドサイトーシスにおいて膜を内側にくぼませる力を提供します。このドメインは膜の特定の曲率を感知し、安定化させる能力を持ち、膜小胞の形成に必要な膜の曲げを促進します。
2. SH3ドメインはプロリン豊富な領域を持つ他のタンパク質との結合に関与しています。このドメインを介して、エンドフィリンはダイナミンやシナプトジャニンなど、膜の切り離しや小胞の成熟に必要な他のタンパク質と相互作用します。ダイナミンは膜を絞り切ることで小胞を細胞膜から分離させるのに必要であり、シナプトジャニンは膜のリサイクルや修復に関与しています。
エンドフィリンはこれらの相互作用を通じて、エンドサイトーシスのさまざまな段階を調整します。特に、エンドフィリンは細胞膜の特定のサイトで膜を曲げること、必要なタンパク質をその場所に集めること、そして最終的には膜を切り離して内部へ物質を取り込む小胞を形成することを助けます。このようにエンドフィリンは、細胞が外部環境から必要な物質を効率的に取り込むための重要な調整者の一つです。
シナプス小胞のリサイクリングとエンドフィリン
シナプス小胞のリサイクリングは、神経伝達物質を含む小胞が神経細胞の末端であるシナプスにおいて放出された後、再び回収されて再利用されるプロセスです。このプロセスは神経伝達の持続性と効率性を保つために重要です。エンドフィリンは、このリサイクリングプロセスにおいて中心的な役割を果たすタンパク質の一つです。
エンドフィリンの機能は、その独特な構造ドメイン、特にSH3ドメインとBARドメインによって支えられています。これらのドメインを通じて、エンドフィリンはシナプス小胞のリサイクリングに関わる以下のような主要な機能を果たします。
1. 膜の曲げとクラスリング: BARドメインを持つエンドフィリンは、膜の曲げを促進し、シナプス小胞の成分を含む膜領域のクラスリング(集合)を助けます。これは、神経伝達物質の放出後に小胞が効率的に回収されるための基盤を作ります。
2. 膜の切り離し: エンドフィリンのSH3ドメインは、ダイナミンなどの他のタンパク質と相互作用し、これにより膜が絞り切られ、小胞が細胞膜から分離されるプロセスが促進されます。これはシナプス小胞が再び充填され、再利用されるための初期ステップです。
3. タンパク質の再配置と小胞の再形成: エンドフィリンはシナプトジャニンとも相互作用し、これによって小胞膜のリサイクルが促進されます。このプロセスには、使用済みの膜のリピッド成分の再構成や、再利用のための小胞の再形成が含まれます。
このようにエンドフィリンは、シナプス小胞のリサイクリングにおける複数の段階で重要な役割を果たし、神経細胞が迅速かつ効率的に神経伝達物質を放出し続けることを可能にします。これは、学習や記憶などの神経系の基本的な機能を支える重要なプロセスです。
第3章:エンドフィリンと神経伝達
シナプス伝達におけるエンドフィリンの重要性
シナプス伝達は、神経細胞間で情報を伝える基本的なプロセスであり、学習、記憶、感覚の処理など、脳の多くの機能に不可欠です。エンドフィリンはこのプロセスにおいて重要な役割を果たします。具体的には、エンドフィリンはシナプス小胞のリサイクリングに関与し、神経伝達物質の効率的なリリースと再利用を支援します。エンドフィリンの主な貢献領域を詳細に見てみましょう。
● シナプス小胞のリサイクリング
– 小胞の回収: 神経伝達物質がシナプス間隙に放出された後、エンドフィリンは使用済みシナプス小胞の膜を細胞内に回収する過程に関与します。このプロセスは、細胞膜の特定の領域での膜の曲げと小胞の形成を含みます。
– ダイナミンとの相互作用: エンドフィリンはダイナミンと結合し、これを活性化します。ダイナミンは、シナプス小胞が細胞膜から切り離されるのを助けるタンパク質であり、この過程はシナプス小胞の回収に不可欠です。
– シナプトタギンとの結合: 一部の研究では、エンドフィリンがシナプトタギンと結合し、シナプス小胞のリリースサイトへの小胞の輸送と再利用を促進する可能性が示唆されています。
● 神経伝達物質のリリースの調整
– 小胞の再充填: 回収されたシナプス小胞は、再び神経伝達物質で充填されます。エンドフィリンはこのプロセスに間接的に関与し、効率的な神経伝達物質のリリースを支えます。
– 膜の再構成: エンドフィリンはシナプトジャニンと協働して膜脂質を再構成し、回収されたシナプス小胞の再形成を支援します。これにより、神経細胞は迅速にシナプス小胞を再利用し、持続的な神経伝達を可能にします。
● シナプス伝達の精度と速度の向上
– シナプス伝達の効率性: エンドフィリンが関与するシナプス小胞の迅速なリサイクリングは、神経伝達の速度と精度を高め、脳の応答性を向上させます。
– 学習と記憶: 効率的なシナプス小胞リサイクリングは、長期的な神経可塑性を支え、学習と記憶形成に重要です。
エンドフィリンのこれらの役割は、シナプス機能の基本的な側面を支え、神経系の健康と疾患において重要です。エンドフィリンの活性が変化することで、シナプス伝達の効率が影響を受け、それが学習障害、記憶喪失、神経変性疾患など、様々な神経系疾患の原因となる可能性があります。実際、アルツハイマー病やパーキンソン病などの条件下では、エンドフィリンの発現や機能が異常を示すことがあり、これらの異常が病態の発展や進行に寄与している可能性が示唆されています。
エンドフィリンは、シナプス機能を正常に維持するための基盤となるタンパク質の一つであり、以下のような複数の重要なプロセスを調整します:
1. 神経伝達物質の効率的な回収とリサイクリング: エンドフィリンは、使用済みのシナプス小胞を効率的に細胞内に回収し、神経伝達物質の再利用を可能にします。これにより、シナプスが迅速に再活性化され、連続した信号伝達が可能になります。
2. シナプス伝達の調整と最適化: エンドフィリンはシナプス伝達の精度を高め、神経細胞間の情報伝達の効率を向上させます。これは、脳の機能を最適化し、学習や記憶などの高次脳機能を支えるために不可欠です。
3. 神経可塑性への寄与: 長期ポテンシャル化(LTP)や長期抑圧(LTD)などの神経可塑性のメカニズムにおいて、エンドフィリンは重要な役割を果たします。これらのプロセスは、学習と記憶において中心的な役割を担っています。
4. 神経系の健康維持と疾患予防: エンドフィリンの正常な機能は、神経系の健康を維持し、神経変性疾患の予防にも寄与する可能性があります。エンドフィリンの機能不全は、シナプス伝達の異常や神経細胞の死に繋がり、神経変性のリスクを高めることが示唆されています。
したがって、エンドフィリンはシナプス伝達の正常な機能に不可欠であり、その機能と調節の理解は、神経系の健康と病態の理解において重要な意味を持ちます。さらに、エンドフィリンを標的とした治療戦略が、将来的には神経変性疾患やその他の神経系疾患の治療に役立つ可能性があります。
シナプス小胞の成長円錐への影響
成長円錐は、神経細胞の軸索の先端に位置する構造で、細胞外環境の化学的な手がかりを感知し、それに応じて軸索の成長方向を決定します。シナプス小胞とそのリサイクリングは、成長円錐の機能と密接に関連しています。エンドフィリンはこのプロセスにおいても重要な役割を果たすと考えられています。具体的には、以下の点で成長円錐への影響が見られます:
● シナプス小胞の供給とリサイクリング
– 成長円錐の動態: 成長円錐が新たなシナプス接続を形成するために前進する際、シナプス小胞は神経伝達物質の放出に必要です。エンドフィリンは、これらの小胞が効率的に輸送され、リサイクリングされることを支援し、成長円錐の活動的な前進を促進します。
– 小胞輸送: シナプス小胞は、成長円錐が細胞外のシグナルに応答して正確に成長するために必要な神経伝達物質やその他の分子を運搬します。エンドフィリンが関与するリサイクリングメカニズムは、これらの小胞が適切に再利用され、円滑に機能することを保証します。
● 神経伝達物質の放出
– 神経伝達物質の局所的な放出: 成長円錐では、シナプス小胞からの神経伝達物質の局所的な放出が、周囲の細胞や組織とのコミュニケーションに重要です。この放出は、成長方向の調節やターゲットとのシナプス形成に影響を与えます。エンドフィリンは、シナプス小胞の輸送、成熟、そして神経伝達物質のリリースプロセスを調節することで、成長円錐の機能を支えます。
● 細胞外シグナルへの応答
– シグナリング経路の調節: エンドフィリンは、細胞外シグナルに対する細胞内応答の調節にも関与しています。成長円錐でのシナプス小胞のリサイクリングとシグナリング分子の放出は、細胞外の環境からの指示に対する適応的な応答を形成します。これにより、成長円錐は環境の化学的勾配を読み取り、軸索の成長を適切な方向に導くことができます。
エンドフィリンを介したこれらのプロセスは、成長円錐の動態と軸索の正確なナビゲーションに不可欠であり、神経回路の形成と神経系の発達において重要な役割を果たします。エンドフィリンの働きによって成長円錐の活動が最適化され、神成長円錐は、神経細胞の軸索の先端に位置する構造で、細胞外環境の化学的な手がかりを感知し、それに応じて軸索の成長方向を決定します。シナプス小胞とそのリサイクリングは、成長円錐の機能と密接に関連しています。エンドフィリンはこのプロセスにおいても重要な役割を果たすと考えられています。具体的には、以下の点で成長円錐への影響が見られます:
● シナプス小胞の供給とリサイクリング
– 成長円錐の動態: 成長円錐が新たなシナプス接続を形成するために前進する際、シナプス小胞は神経伝達物質の放出に必要です。エンドフィリンは、これらの小胞が効率的に輸送され、リサイクリングされることを支援し、成長円錐の活動的な前進を促進します。
– 小胞輸送: シナプス小胞は、成長円錐が細胞外のシグナルに応答して正確に成長するために必要な神経伝達物質やその他の分子を運搬します。エンドフィリンが関与するリサイクリングメカニズムは、これらの小胞が適切に再利用され、円滑に機能することを保証します。
● 神経伝達物質の放出
– 神経伝達物質の局所的な放出: 成長円錐では、シナプス小胞からの神経伝達物質の局所的な放出が、周囲の細胞や組織とのコミュニケーションに重要です。この放出は、成長方向の調節やターゲットとのシナプス形成に影響を与えます。エンドフィリンは、シナプス小胞の輸送、成熟、そして神経伝達物質のリリースプロセスを調節することで、成長円錐の機能を支えます。
● 細胞外シグナルへの応答
– シグナリング経路の調節: エンドフィリンは、細胞外シグナルに対する細胞内応答の調節にも関与しています。成長円錐でのシナプス小胞のリサイクリングとシグナリング分子の放出は、細胞外の環境からの指示に対する適応的な応答を形成します。これにより、成長円錐は環境の化学的勾配を読み取り、軸索の成長を適切な方向に導くことができます。
エンドフィリンを介したこれらのプロセスは、成長円錐の動態と軸索の正確なナビゲーションに不可欠であり、神経回路の形成と神経系の発達において重要な役割を果たします。エンドフィリンの働きによって成長円錐の活動が最適化され、神
第4章:エンドフィリンの細胞生物学的研究
細胞内でのエンドフィリンの動態と観察技術
エンドフィリンの細胞内での動態は、細胞の生理機能、特にエンドサイトーシスやシナプス小胞のリサイクリングにおいて重要な役割を果たします。この動態を理解するためには、細胞生物学および分子生物学の高度な観察技術が必要です。以下は、エンドフィリンの動態を観察するために用いられる主な技術です。
● フルオレセンス顕微鏡法
– フルオレセントプローブ: エンドフィリンに特異的なフルオレセンスタグ(例えば、GFP(緑色蛍光タンパク質)やmCherry)を組み込むことで、生きた細胞内でのエンドフィリンの局在や動態をリアルタイムで観察できます。
– FRAP(Fluorescence Recovery After Photobleaching): フルオレセンス分子を特定の領域で一時的に消失させた後の回復プロセスを追跡することで、タンパク質の動態や細胞内での拡散速度を評価できます。
● 共焦点レーザー顕微鏡法
共焦点レーザー顕微鏡を使用することで、細胞内の厚いセクションを高解像度で観察し、エンドフィリンの正確な細胞内局在を三次元的に解析することが可能になります。
● 電子顕微鏡(EM)
電子顕微鏡法は、細胞内のエンドフィリンの局在を超高解像度で観察するのに用いられます。特に、イムノゴールドラベリングを利用すると、特定のタンパク質に対する抗体に金粒子を結合させ、そのタンパク質の細胞内での正確な位置を電子密度の高いマーカーとして可視化できます。
● タイムラプスイメージング
生細胞におけるエンドフィリンの動的な挙動を時間経過で追跡するために、タイムラプスイメージングが有効です。これにより、エンドサイトーシスやシナプス小胞のリサイクリングなどのプロセス中でのエンドフィリンの役割を動的に観察できます。
● FRET(Förster Resonance Energy Transfer)
FRETは、二つのフルオロフォア間のエネルギー移動を利用して、タンパク質間の相互作用やその動的な変化を検出する技術です。エンドフィリンが他のタンパク質と相互作用する際の動態を解析するのに適しています。
これらの技術によって、エンドフィリンの細胞内での動態やその生理的機能に対する理解が深まります。例えば、エンドフィリンが細胞膜の特定の領域にどのようにリクルートされ、エンドサイトーシスや小胞輸送にどのように関与するか、さらにはエンドフィリンの機能異常が細胞の挙動や病態にどのように影響するかといった情報が明らかになります。これは、エンドフィリンが細胞内で果たす役割を正確に理解し、その機能を標的とした新たな治療戦略を開発する上で非常に重要です。
たとえば、エンドフィリンの活性が変化すると、神経伝達物質のリリースやリサイクリングが不適切になり、神経系の疾患や発達障害につながる可能性があります。また、がん細胞におけるエンドサイトーシスの異常は、細胞増殖や転移に影響を与えることが知られています。したがって、エンドフィリンの正確な動態を観察し、その調節機構を理解することは、これらの疾患のメカニズムを解明し、新しい治療法の開発に繋がります。
さらに、エンドフィリンの動態を細かく観察する技術は、細胞生物学の基礎研究だけでなく、薬理学的研究においても応用されます。特定の薬剤がエンドフィリンの活性にどのように影響するか、またはエンドフィリンを介したシグナル伝達経路をどのように調節するかを理解することで、より効果的な薬剤のスクリーニングや創薬への道が開けます。
最終的に、これらの高度な観察技術による研究成果は、エンドフィリンというタンパク質が細胞機能に及ぼす広範な影響を理解することを可能にし、生命科学の多様な分野での新たな発見へとつながるでしょう。エンドフィリンに関するさらなる研究は、基礎生物学から臨床応用に至るまで、多くの重要な洞察を提供することが期待されています。
エンドフィリンとクラスリンの相互作用
エンドフィリンとクラスリンはエンドサイトーシスにおいて密接に協力し合うタンパク質です。エンドサイトーシスは細胞がその膜を内側にくぼませ、細胞外から物質を取り込むプロセスであり、細胞の栄養摂取、シグナル伝達、および細胞表面の受容体の調節に不可欠です。
● エンドフィリンとクラスリンの機能
– エンドフィリンは、細胞膜の特定のリゾン(取り込みを促進する膜領域)で膜を曲げることにより、エンドサイトーシスを促進します。エンドフィリンは、そのBARドメインを介して膜を曲げる能力があり、SH3ドメインを介して、ダイナミンのような他のエンドサイトーシスに関わるタンパク質と相互作用します。
– クラスリンは、エンドサイトーシスにおける別の重要なタンパク質で、トリスケリオンと呼ばれる構造を形成します。これは、細胞膜にクラスリン被覆小胞を形成することにより、取り込みプロセスを助けます。クラスリン被覆小胞は、細胞内に物質を取り込むための一次的な手段です。
● エンドフィリンとクラスリンの相互作用
エンドフィリンとクラスリンの相互作用は、エンドサイトーシスプロセスの初期段階において重要です。エンドフィリンは、クラスリン被覆小胞の形成を促進し、また、ダイナミンと共にクラスリン被覆小胞の成熟と細胞膜からの切り離しを助けることにより、効率的な物質の取り込みを支援します。具体的には、エンドフィリンは次のような役割を果たします:
1. 膜の曲げ: エンドフィリンのBARドメインは、細胞膜の特定の領域を曲げることによって、クラスリン被覆小胞の形成を促進します。
2. タンパク質のリクルート: エンドフィリンのSH3ドメインは、ダイナミンやシナプトジャニンなどの他のエンドサイトーシス関連タンパク質を膜へリクルートし、これらのタンパク質がクラスリン被覆小胞の成熟と切り離しに必要です。
3. 相互作用の調節: エンドフィリンとクラスリン(及びその他のタンパク質)間の相互作用は、細胞の状態や取り込まれる物質の種類に応じて調節されます。この調節により、細胞は外部環境からのさまざまなシグナルに適切に反応することができます。
エンドフィリンとクラスリンの相互作用の調節は、エンドサイトーシスプロセスの効率と特異性を高めることに寄与します。この相互作用は、特定の受容体やリガンドが細胞内に効果的に取り込まれることを保証し、細胞の応答性や生存に不可欠な多くのシグナル伝達経路の調節に影響を与えます。
● 相互作用の詳細なメカニズム
1. 膜の特異的な認識: エンドフィリンは特定のリゾンを認識し、そこにクラスリンと他のエンドサイトーシス関連タンパク質を集めることで、小胞の形成を促進します。このプロセスは、膜受容体が特定のリガンドに結合した後に特に活発化します。
2. 小胞の成熟と切り離し: クラスリン被覆小胞が形成されると、エンドフィリンはダイナミンと共に作用し、細胞膜から小胞を切り離します。この段階では、エンドフィリンは小胞の脱被覆(クラスリンの除去)を促進する役割も果たす可能性があり、これにより小胞が細胞質に放出され、取り込まれた物質が細胞内で利用される準備が整います。
3. 細胞内シグナリングの調節: エンドフィリンとクラスリンの相互作用は、細胞内シグナリング経路にも影響を及ぼします。例えば、成長因子受容体やその他のシグナル分子の取り込みは、細胞の成長、分化、生存の決定に直接関与します。エンドフィリンとクラスリンによる効率的なエンドサイトーシスは、これらのシグナル伝達経路のダイナミクスを調節し、細胞応答を最適化します。
● 研究と臨床への応用
エンドフィリンとクラスリンの相互作用を理解することは、細胞生物学だけでなく、疾患の治療戦略の開発にも応用可能です。例えば、がん細胞ではエンドサイトーシスが異常に活性化されることがあり、これが細胞の成長や転移に寄与していることが知られています。エンドフィリンとクラスリンの活動を特異的に調節することで、がん細胞の成長を抑制する新たな治療法の開発が期待されます。
また、アルツハイマー病やパーキンソン病などの神経変性疾患において、エンドサイトーシスプロセスの異常が病態に関与していることが示唆されています。エンドフィリンとクラスリンの相互作用の詳細な理解は、これらの疾患の病態メカニズムを解明し、新たな治療標的を特定する手がかりを提供する可能性があります。具体的には、エンドサイトーシスの異常が神経細胞の機能障害や細胞死に直接関与している場合、エンドフィリンとクラスリンの活性を正常化することで、これらの疾患の進行を遅らせるかもしれません。
さらに、エンドフィリンとクラスリンの相互作用の調節は、薬剤送達システムの設計にも応用できます。エンドサイトーシスを利用した薬剤送達は、特定の細胞や組織に薬剤を効率的に届ける有望な手法です。エンドフィリンとクラスリンの活動をコントロールすることで、薬剤が目的の細胞に取り込まれやすくなり、治療効果を高めることができるでしょう。
このように、エンドフィリンとクラスリンの相互作用に関する研究は、細胞生物学の基礎研究から臨床応用に至るまで、幅広い分野での新たな可能性を開くことが期待されます。疾患治療における新しいアプローチの開発だけでなく、細胞の基本的な機能理解の向上にも貢献するでしょう。今後、エンドフィリンとクラスリンの相互作用をさらに詳細に理解するための研究が進むことで、これらの可能性が現実のものとなることが期待されます。
第5章:エンドフィリン関連疾患と治療への応用
エンドフィリンの異常は、細胞内のエンドサイトーシスプロセスの障害につながり、多様な疾患の発症に関与しています。エンドフィリンは細胞膜の形状変化、特定の受容体の取り込みとリサイクリング、シナプス小胞の再利用など、細胞内で幅広い役割を果たします。エンドフィリンの機能不全は、以下のような疾患や状態と関連していることが知られています。
● 神経変性疾患
– アルツハイマー病: アルツハイマー病の患者では、エンドサイトーシスの異常が報告されており、これがアミロイドβペプチドの異常な蓄積に寄与している可能性があります。エンドフィリンの機能不全は、細胞内でのアミロイド前駆体タンパク質(APP)の処理に影響を及ぼし、アミロイドβの生成を促進する可能性があります。
– パーキンソン病: パーキンソン病では、α-シヌクレインというタンパク質の凝集が主な特徴の一つです。エンドフィリンの異常は、α-シヌクレインの凝集と細胞への取り込みに影響を与え、神経細胞の機能障害や細胞死を引き起こす可能性があります。
● 癌
– がん細胞の増殖と転移: エンドフィリンの異常は、がん細胞の増殖、生存、転移に関わるシグナル伝達経路の調節に影響を及ぼします。特に、エンドサイトーシスを介した成長因子受容体のリサイクリングの異常は、がん細胞の増殖を促進する可能性があります。
● 精神疾患
– 双極性障害、統合失調症: エンドフィリンの遺伝的変異は、双極性障害や統合失調症などの精神疾患のリスクと関連しています。エンドフィリンが関与する神経伝達物質の再取り込みやシナプス機能の障害は、これらの疾患の発症機序に影響を与える可能性があります。
● 心血管疾患
エンドフィリンは血管内皮細胞の機能にも影響を及ぼし、その異常は血管の新生や修復過程に影響を与える可能性があります。これにより、動脈硬化や他の心血管疾患のリスクが高まるかもしれません。
● エンドフィリンの異常を標的とした治療戦略
エンドフィリンの異常に基づく疾患の治療戦略には、エンドフィリンの機能を正常化する方法が含まれます。これには、エンドフィリンの過剰な活性を抑制するか、不足している機能を補充するアプローチがあります。具体的な治療戦略としては以下のような方法が考えられます。
1. 小分子化合物: エンドフィリンの活性を特異的に調節する小分子化合物の開発は、エンドフィリンの機能異常に関連する疾患の治療に有望です。これらの化合物は、エンドフィリンの特定のドメインと相互作用し、その活性を調節することで、エンドサイトーシスのプロセスを正常化させることが目指されます。
2. 遺伝子療法: エンドフィリンの遺伝的変異が疾患の原因である場合、遺伝子療法を用いて正常なエンドフィリンのコピーを細胞に導入することで、異常なシグナル伝達や細胞機能を修正することができます。このアプローチは、特に遺伝性疾患の治療に有効です。
3. RNA干渉技術: エンドフィリンの過剰発現が問題となる病態には、RNA干渉技術を用いてエンドフィリンmRNAのレベルを特異的に低下させることで、タンパク質の発現を抑制することが可能です。これにより、エンドフィリンの過剰な活性を抑え、細胞機能を正常化させることができます。
4. 抗体療法: エンドフィリンやその相互作用パートナーに対する特異的な抗体を開発し、これを用いてエンドフィリンの活性を調節する方法も考えられます。この方法は、エンドフィリンが過剰に活性化している状態や、特定の相互作用を阻害することが治療的に有益である場合に有効です。
これらの治療戦略の開発には、エンドフィリンの正確な生物学的役割と疾患における具体的な機能異常を理解することが不可欠です。現在のところ、多くの研究が進行中であり、エンドフィリンに関連する疾患の治療法の改善に向けた新たな知見が期待されています。これらの治療法が実際に臨床で使用されるには、さらなる研究と試験が必要ですが、エンドフィリンを標的としたアプローチは、多くの疾患の治療に新たな展望を開く可能性を持っています。







