異数性とは、細胞の染色体数が正常な個体のそれと異なる状態を指します。この変化は、生物の成長や発達に大きな影響を及ぼし、特に人間においては、ダウン症やターナー症といった特定の疾患の原因となります。異数性は、染色体の分配が細胞分裂時に不均等に行われることによって発生します。この染色体の数の変化は、遺伝情報の不均衡を引き起こし、発達遅延、身体的特徴の変化、生殖能力の問題など、多岐にわたる影響をもたらす可能性があります。
染色体異常とは何か
染色体異常とは、染色体の数や構造に生じた変化により、個体の発達、健康、または生殖能力に影響を与える遺伝的障害です。これらの異常は、染色体の欠損、重複、転座、または数の変化など様々な形で現れ、個人の遺伝的特性や疾病のリスクに大きく影響を及ぼします。
染色体の基本構造と機能
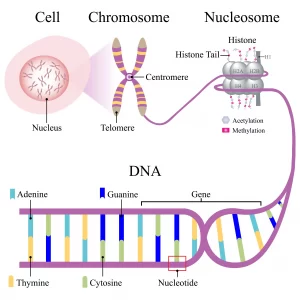
染色体は、DNAとタンパク質から成る複雑な構造体であり、生物の遺伝情報が格納されています。DNAは、遺伝子の形で特定の遺伝情報をコードしており、タンパク質はDNAの構造を維持し、遺伝子の発現を制御する役割を担います。染色体の構造と機能について、以下に基本を説明します。
### 染色体の基本構造
– DNA: 長い二重螺旋構造をしており、遺伝情報をコードする化学物質です。
– ヒストン: DNAが巻きついているタンパク質。DNAをコンパクトに折りたたんで染色体を形成し、遺伝子のアクセス性を調節します。
– クロマチン: DNAとヒストンが複合した構造。クロマチンの緊密度は遺伝子の発現に影響を与えます。
– セントロメア: 染色体の中央部に位置し、細胞分裂時に姉妹染色分体をつなぎ止める役割を持ちます。
– テロメア: 染色体の端部に位置し、染色体の安定性を保持する重要な役割を果たします。
### 染色体の機能
– 遺伝情報の保存: 生物の遺伝情報はDNA上の遺伝子によってコードされ、染色体内に安全に保存されます。
– 遺伝情報の伝達: 細胞分裂時に染色体は複製され、親細胞から娘細胞へ正確に遺伝情報が伝達されます。
– 遺伝子の発現調節: 染色体上の遺伝子は、生物の発達や環境への応答に応じて発現されます。クロマチンの構造変化により、遺伝子のアクセス性が調節され、遺伝子発現が制御されます。
– 遺伝的多様性の生成: 有性生殖において、親から子へ遺伝情報が伝達される過程で、遺伝的再組換えが発生し、多様性が生み出されます。
染色体は、これらの基本構造と機能を通じて、生物の遺伝的特性を決定し、種の継続と進化に重要な役割を果たしています。
異数性を含む染色体異常の種類
異数性を含む染色体異常は、染色体の数に関する異常であり、主に二つの大きなカテゴリーに分けられます。これらの異常は、遺伝的疾患の原因となることが多く、個体の発達、健康、生殖能力に影響を及ぼすことがあります。
### 1. アネプロイディaneuploidy
アネプロイディは、特定の染色体が正常な数よりも多いか少ない状態を指します。これは、細胞分裂時の染色体の不正確な分配によって発生します。アネプロイディにはいくつかの一般的な形態があります。
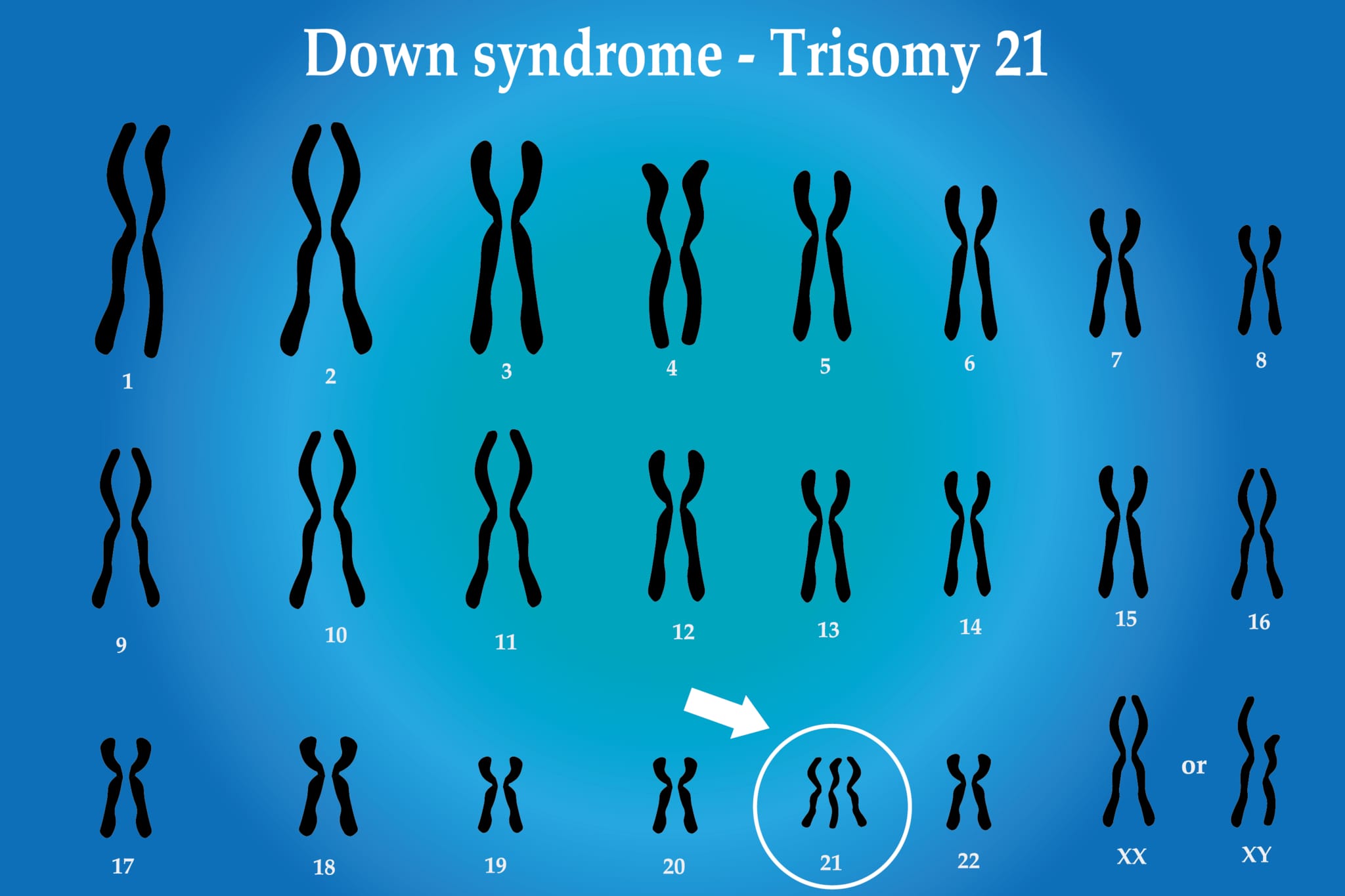
– トリソミー: 特定の染色体が1本余分に存在する状態。例えば、ダウン症候群(21トリソミー)は、21番染色体が3本あることによって特徴付けられます。
– モノソミー: 特定の染色体が1本しかない状態。ターナー症候群は、女性がX染色体を1つしか持たない状態であり、これが一例です。
### 2. ポリプロイディ
ポリプロイディは、染色体の一式(ハプロイド数)が正常よりも一つ以上多い状態を指します。これは、通常、全ての染色体が影響を受けます。
– トリプロイディ(三倍体): 細胞が3セットの染色体を持つ状態(通常は2セット)。これは、生物全体では稀ですが、一部の植物種や昆虫では一般的に見られます。
– テトラプロイディ(四倍体): 細胞が4セットの染色体を持つ状態。これも特定の生物で見られる現象です。
これらの染色体異常は、遺伝的検査によって診断され、個々の異常に応じた管理や支援が必要となることがあります。異数性を含む染色体異常は、遺伝的カウンセリングの重要なトピックであり、特に妊娠中や遺伝病のリスクが高い家族において重要です。
異数性の原因と発生メカニズム
細胞分裂のエラーと異数性の関連性
細胞分裂のエラーは異数性の主な原因の一つです。細胞分裂、特に減数分裂の過程で正確な染色体の分配が行われない場合、異数性が生じる可能性があります。異数性は、特定の染色体が正常な数よりも多いか少ない状態を指し、このような染色体数の不均衡は、多くの場合、細胞分裂時のエラーに起因します。
### 細胞分裂と異数性の関連性
1. 減数分裂中の非分離(Nondisjunction):
減数分裂は、生殖細胞(精子や卵子)を生成する過程です。この過程では、染色体は通常、2回の細胞分裂を経て、親細胞の染色体数の半分を含む生殖細胞を形成します。しかし、第一減数分裂または第二減数分裂の際に染色体が正しく分離しない非分離が発生すると、異数性が生じる原因となります。たとえば、一方の生殖細胞が余分な染色体を持ち、もう一方がその染色体を欠く場合、受精した後の胚にトリソミーやモノソミーといった異数性状態が生じます。
2. 有糸分裂中のクロマチド非分離:
有糸分裂は体細胞が分裂する過程で、通常、染色体は姉妹クロマチドに分かれ、それぞれが新しい細胞に均等に分配されます。しかし、クロマチド(染色体の複製コピー、染色分体)が正常に分離しない場合(クロマチド非分離)、細胞は正常な染色体数より多いか少ない染色体を持つことになり、これが体細胞の異数性につながります。
### 異数性の影響
異数性は発達障害、遺伝性疾患、不妊症など、さまざまな健康上の問題を引き起こす可能性があります。例えば、トリソミー21(ダウン症候群)は、21番染色体が1本余分に存在する状態であり、特定の身体的特徴や学習障害を引き起こします。
細胞分裂のエラーによる異数性の発生は、生物学的プロセスの複雑さを示すとともに、遺伝学と医学研究における重要な研究領域です。これらのエラーの理解と、それによって引き起こされる条件の管理や治療への知見は、遺伝病の診断や治療法の開発に不可欠です。
胚発生における異数性の影響
胚発生における異数性は、発達の初期段階で重要な影響を及ぼす可能性があります。異数性、すなわち染色体の数に関する異常は、胚の成長、分化、および正常な器官形成において重要な役割を果たす遺伝子の機能に影響を与えることがあります。これにより、発達障害、先天性異常、または妊娠の早期段階での流産が引き起こされることがあります。
### 胚発生への影響
1. 発達障害と先天性異常:
異数性は、特定の染色体上に位置する遺伝子のコピー数を変更するため、胚が正常に発達するために必要な精密な遺伝的調節を乱す可能性があります。例えば、ダウン症候群(21トリソミー)の場合、余分な21番染色体によって遺伝子の表現が変化し、知的障害や心臓疾患などの先天性異常を引き起こすことが知られています。
2. 妊娠の損失:
胚発生初期における異数性は、しばしば妊娠の損失につながります。多くの場合、異数性を持つ胚は、自然な選択の過程で早期段階で流産することが多く、これは自然流産の一般的な原因の一つです。特に、重大な異数性(例えば、多くの染色体に影響を与えるもの)は、胚の生存に不可欠な遺伝子の機能に重大な影響を及ぼす可能性があります。
3. 不妊症:
また、異数性は不妊症の原因となることもあります。特に、生殖細胞(精子や卵子)における異数性は、受精能力の低下や受精後の胚の発達障害につながる可能性があります。
### 管理と介入
胚発生における異数性の影響は深刻であることが多いため、先天性異常や遺伝病のリスクが高い妊娠においては、遺伝学的スクリーニングや診断が推奨されます。これには、胎児の染色体異常を検出するための羊水検査や絨毛検査などが含まれます。また、体外受精(IVF)を通じての先発遺伝学的診断(PGD)は、異数性リスクが高い場合に、正常な染色体数を持つ胚を選択するために使用されることがあります。
胚発生における異数性の理解は、遺伝病や発達障害の早期診断と管理に不可欠であり、遺伝学、胚発生学、および産科学の分野における継続的な研究の対象となっています。
異数性の検査方法とその重要性
PGTとは:胚の染色体スクリーニング
PGT(Preimplantation Genetic Testing)は、体外受精(IVF)によって得られた胚の遺伝的スクリーニングを行う技術です。この検査は、胚を母体に移植する前に、遺伝的異常がないかを調べることを目的としています。PGTには主に3つのタイプがあり、それぞれ異なる遺伝的状態を検出することができます。
### PGTのタイプ
1. PGT-A(PGT for Aneuploidies):
PGT-Aは、胚の染色体異数性を検査するもので、特定の染色体が正常な数よりも多いか少ないかを調べます。これにより、ダウン症候群(21トリソミー)やターナー症候群(Xモノソミー)など、染色体異数性に関連する遺伝的状態のリスクを持つ胚を特定することができます。
2. PGT-M(PGT for Monogenic/Single Gene Disorders):
PGT-Mは、単一遺伝子疾患を持つ胚を検出するための検査です。これは、ハンチントン病や嚢胞性線維症など、特定の遺伝子変異によって引き起こされる遺伝性疾患を対象としています。この検査により、特定の遺伝病を発症するリスクが高い胚を事前に識別することが可能です。
3. PGT-SR(PGT for Structural Rearrangements):
PGT-SRは、染色体の構造的再編成を持つ胚を識別する検査です。これには、染色体の転座や欠失などが含まれます。この検査は、特定の染色体構造異常を伴う遺伝的疾患のリスクがある家族で使用されることがあります。
### PGTの意義
PGTは、遺伝的リスクが高いカップルにとって貴重な選択肢を提供します。この技術により、遺伝的疾患のリスクを持つ胚が母体に移植される前に特定され、健康な胚を選択することが可能となります。これにより、遺伝的疾患を持つ子どもが生まれるリスクを軽減し、不妊治療を受けるカップルに希望をもたらします。
また、PGTは不妊治療の成功率を高めることにも寄与することが示されています。特に高齢の母親や遺伝的リスクを持つカップルでは、健康な胚を選択することで、妊娠の成功率と出生後の健康な成果を向上させることができます。
次世代シーケンサによる詳細な解析
次世代シーケンサ(Next-Generation Sequencing, NGS)技術を用いることにより、染色体の異数性や遺伝子の変異を高精度で詳細に解析することが可能です。この技術は、遺伝学、がん研究、疾患の診断、および個別化医療に革命をもたらしています。NGSを用いた異数性の解析は、遺伝的多様性を理解し、特定の疾患や個体差に関連する遺伝的要因を明らかにする上で重要な役割を果たします。
### 異数性のNGSによる解析
1. 高スループット解析:
NGSは、同時に数百万から数十億のDNA断片をシーケンスすることができる高スループット技術です。これにより、全ゲノムや特定の遺伝子領域の詳細な解析が可能となり、染色体異数性を含む遺伝的変異を迅速かつ大規模に同定できます。
2. 異数性の正確な同定:
NGSによる解析は、染色体のコピー数変異(CNV)や微細な遺伝子変異を含む異数性の特定に非常に有効です。これにより、疾患の原因となる染色体異常や、遺伝性疾患のリスク要因を正確に識別することが可能になります。
3. 包括的な遺伝的プロファイリング:
NGSを用いた包括的な遺伝的プロファイリングにより、個人の遺伝的リスクや疾患の傾向を詳細に評価できます。これは、個別化医療や精密医療の実現に向けた重要なステップです。
4. 新たな遺伝的変異の発見:
NGSは、未知の遺伝的変異や異常な遺伝子の組み合わせを発見する能力を持っています。これにより、新たな疾患関連遺伝子の同定や、遺伝子機能の理解が深まります。
### 応用分野
– がん遺伝学: がん細胞における染色体異数性や遺伝子変異を解析し、がんの発生メカニズムを理解し、ターゲット治療の選択を支援します。
– 遺伝性疾患の診断: 遺伝性疾患やレア病の原因となる遺伝子変異を同定し、早期診断や適切な治療戦略の決定に役立てます。
– 生殖医療: IVFにおける胚の遺伝的スクリーニングにNGSを利用し、遺伝的リスクを持つ胚の選択を支援します。
NGSによる異数性の詳細な解析は、遺伝的疾患の理解と管理を向上させ、個別化された治療アプローチの開発に貢献しています。この技術の進展により、遺伝学研究の新たな地平が開かれつつあります。
異数性に関連する疾患と症状
一般的な異数性疾患の例
異数性疾患は、染色体の数の異常によって引き起こされる遺伝性の状態です。これらの疾患は、特定の染色体が正常よりも多いか少ないことに起因し、多岐にわたる身体的特徴や発達障害を引き起こすことがあります。以下は、異数性に関連する一般的な疾患の例です。
### ダウン症候群(トリソミー21)
ダウン症候群は、21番染色体が3本存在することによって引き起こされる最も一般的な染色体異常の一つです。この追加の染色体が引き起こす影響には、知的障害、特徴的な顔貌、および心臓疾患が含まれます。
### エドワーズ症候群(トリソミー18)
エドワーズ症候群は、18番染色体の異数性によって特徴づけられます。この状態は、重度の発達障害、成長遅延、および多くの場合、生後早期に致命的である多数の身体的異常を引き起こします。
### パトー症候群(トリソミー13)
パトー症候群は、13番染色体の異数性に関連しています。この疾患は、重度の知的障害、成長障害、および様々な先天性異常を引き起こすことが知られています。生存率は低く、多くの患者は生後数ヶ月以内に亡くなります。
### ターナー症候群
ターナー症候群は、女性がX染色体を1つしか持たないか、もしくはX染色体が部分的に欠如している状態です。この疾患は、成長障害、不妊、心臓疾患などを引き起こす可能性があります。
### クラインフェルター症候群
クラインフェルター症候群は、男性が余分なX染色体を持つことにより発生します(通常はXXY)。この状態は、性的発達の遅延、低いテストステロンレベル、および不妊を引き起こすことがあります。
これらの異数性疾患は、遺伝的検査によって診断されることが多く、個々の疾患に応じた治療やサポートが提供されます。遺伝的カウンセリングは、これらの疾患を持つ個人やその家族にとって重要なリソースとなります。
異数性がもたらす健康への影響
異数性は、染色体の数に関する異常であり、人の健康に多岐にわたる影響を及ぼすことがあります。異数性が原因で発生する健康上の問題は、軽度から重度に至るまでさまざまです。異数性がもたらす主な健康への影響を以下に示します。
### 発達障害
異数性を持つ多くの人々は、学習困難や知的障害を経験します。これは、特定の染色体異常が脳の発達に影響を与えるためです。例えば、ダウン症候群(トリソミー21)の患者は、一般的に軽度から中度の知的障害を持つことが知られています。
### 身体的特徴
異数性は、特有の身体的特徴を引き起こすことがあります。例として、ダウン症候群の患者は、斜めに上向きの目や小さな手足を持つことがあり、ターナー症候群の女性は、低身長や翼状頸(首の側面に余分な皮膚がある状態)を持つことがあります。
### 生殖健康への影響
特定の異数性状態は、生殖機能に影響を与えることがあります。例えば、クラインフェルター症候群(XXY)の男性は、しばしば不妊症に悩まされます。ターナー症候群の女性も、不妊の問題に直面することがあります。
### 心臓病とその他の健康問題
異数性は、心臓病や先天性心疾患のリスクを高めることがあります。ダウン症候群の個体は、心臓に関連する健康問題を抱えることが一般的です。また、異数性を持つ人々は、聴覚障害や免疫系の問題など、他の健康上の問題を経験することがあります。
### 早期の介入と管理
異数性がもたらす健康上の問題に対処するためには、早期の介入と適切な医療管理が重要です。これには、特定の状態に対応した教育プログラム、身体的な治療、および必要に応じた医療処置が含まれます。遺伝的カウンセリングは、異数性疾患を持つ人々とその家族にとって貴重なサポートを提供し、疾患の理解と管理に役立ちます。
異数性によってもたらされる健康への影響は、個人によって大きく異なるため、個々のニーズに合わせたケアが重要です。専門家による継続的なサポートとケアを受けることで、異数性を持つ人々も健康で充実した生活を送ることができます。
異数性診断後の医療的対応
胚移植と異数性:リスクとメリット
胚移植は不妊治療の一環として行われる体外受精(IVF)のプロセスで、受精卵(胚)を女性の子宮内に移植する手法です。異数性、つまり染色体の数に関する異常は、胚に影響を及ぼす可能性があり、このプロセスにおいて重要な考慮事項となります。胚移植と異数性に関連するリスクとメリットを理解することは、患者が適切な意思決定を行う上で重要です。
### リスク
1. 異数性胚の移植リスク:
異数性を持つ胚を移植すると、妊娠に至らない可能性が高くなるほか、妊娠しても流産のリスクが増加します。また、生まれてくる子供が染色体異常を持つリスクも高まります。通常の体外受精で産生された胚と比較して、ICSI(顕微授精)で産生された胚の方が異数性であることが多いことが示されています。(ソース)
2. 染色体異常を持つ子供の出産:
異数性を持つ胚が正常に成長した場合、ダウン症候群やエドワーズ症候群などの染色体異常を持つ子供が生まれる可能性があります。これらの状態は、様々な健康問題や発達障害を引き起こす可能性があります。
### メリット
1. 異数性のスクリーニングと選択:
着床前遺伝子診断(PGD)や包括的染色体スクリーニング(PGS/PGT-A)などの技術を用いて、移植前に胚の染色体を分析することが可能です。これにより、染色体異常のない胚を選択し、妊娠の成功率を高めることができます。
2. 遺伝病のリスク軽減:
特定の遺伝病を持つ家族歴がある場合、PGDを使用してその病気の遺伝子変異を持つ胚を特定し、避けることができます。これにより、遺伝病のリスクを持つ子供が生まれる確率を大幅に低減させることが可能です。
3. 不妊治療の成功率向上:
染色体異常のない胚のみを選択することで、不妊治療の成功率を向上させることができます。これは特に高齢の女性や、遺伝的リスクを持つカップルにとって有益です。
胚移植と異数性に関するリスクとメリットを十分に理解し、適切なスクリーニングやカウンセリングを受けることで、患者は自身の状況に最適な選択を行うことができます。異数性のリスクを管理し、健康な妊娠を目指すためには、医療提供者と密接に協力することが重要です。
遺伝カウンセリングの役割
異数性に関する遺伝カウンセリングは、染色体異常のリスクが懸念される個人や家族に対して重要なサポートを提供します。染色体の数に関する異常は、発達障害、身体的特徴、繁殖能力の問題など、多様な健康上の影響を及ぼす可能性があるため、遺伝カウンセリングの役割は多岐にわたります。
### 情報提供
臨床遺伝専門医(遺伝カウンセラー)は、異数性の基礎知識、それがもたらす可能性のある健康上の影響、そして利用可能な診断手段や治療オプションについて、詳細かつ正確な情報を提供します。また、特定の異数性状態に関連する遺伝的リスクや疾患の発生確率についても説明します。
### リスク評価
遺伝カウンセリングは、個人またはその家族が特定の染色体異常を持つ子を持つリスクを評価するのに役立ちます。家族歴や既存の医療情報を基に、遺伝的リスクを評価し、必要に応じて遺伝子検査や染色体分析を勧めることがあります。
### サポートとガイダンス
異数性の診断を受けた個人や家族に対して、遺伝カウンセラーは心理的サポートを提供し、将来に向けての計画を立てる際のガイダンスを提供します。これには、治療選択肢の検討、特別な教育やサポートサービスへのアクセス、さらには家族計画のアドバイスも含まれます。
### 意思決定のサポート
遺伝カウンセリングは、遺伝的リスクに基づいた意思決定をサポートします。これには、遺伝子検査を受けるかどうか、または不妊治療のオプション(例:体外受精におけるPGT)を検討する際の意思決定が含まれます。カウンセラーは、患者が情報に基づいた選択を行えるよう支援します。
### 教育と啓発
臨床遺伝専門医(遺伝カウンセラー)は、異数性および関連する遺伝的状態について、患者やその家族だけでなく、一般の人々や医療提供者に対しても教育と啓発を行います。これにより、社会全体の理解を深め、関連する遺伝的状態に対する誤解を解消することができます。
遺伝カウンセリングは、異数性に直面している個人や家族に対して、情報、支援、そして希望を提供する重要なサービスです。専門的な知識と共感的なサポートを通じて、遺伝カウンセラーは、患者が自身の遺伝的健康に対する理解を深め、ポジティブな生活計画を立てるのを助けます。
異数性の研究と最新情報
異数性研究の現状と未来
異数性に関する研究は、医学と遺伝学の分野で継続的に進展しています。これらの研究は、染色体の数に関する異常とそれが人の健康に与える影響をより深く理解することを目的としています。現状と将来の見通しを以下に概説します。
### 現状
– 遺伝子検査の進化: 次世代シーケンシング(NGS)技術の発展により、異数性を含む遺伝的状態の診断がより迅速かつコスト効率良く可能になりました。これにより、疾患の原因を特定し、遺伝的リスクの評価が容易になっています。
– 疾患メカニズムの理解: 異数性に関連する特定の疾患のメカニズムについての研究が進行中です。これは、新しい治療法や管理戦略の開発に役立つ可能性があります。
– 個別化医療への貢献: 異数性の精密な診断は、より効果的な治療法や個別化された医療介入の選択を可能にします。
### 未来
– 治療法の革新: CRISPR-Cas9などのゲノム編集技術の進歩により、将来的には異数性に関連する遺伝子異常を修正することが可能になるかもしれません。これは、遺伝性疾患の根本的な治療への道を開くことが期待されています。
– 胚のスクリーニング技術の改善: IVFにおける胚の選択プロセスがさらに進化することで、健康な妊娠の確率を高めることができます。先発遺伝子診断(PGD)や包括的染色体スクリーニング(PGT-A)などの技術がさらに洗練され、利用可能になるでしょう。
– 予防戦略の開発: 異数性に関する深い理解は、特定の染色体異常の予防に向けた戦略を開発する上で重要です。ライフスタイルの変更や特定の医療介入により、異数性リスクを低減することが可能になるかもしれません。
– 公衆衛生への影響: 異数性に関する一般の認識の向上と遺伝カウンセリングの普及により、遺伝性疾患の予防と管理に対する公衆衛生アプローチが強化されます。
異数性研究の未来は、遺伝学、分子生物学、医療技術の進歩によって形作られます。これらの進展は、異数性に関連する疾患の診断、治療、予防の方法を根本的に変える可能性があり、患者の生活の質の向上に大きく貢献することが期待されています。
2022年からの新しい発見と進歩
異数性の研究は、近年大きく進歩しており、特に母系のゲノムにおける異数性の原因や影響に関する理解が深まっています。例えば、人間の妊娠において、多くの受精卵が胚盤胞へと発達せず、また胚盤胞が形成されたとしても、着床に至らない場合があります。妊娠初期の流産の最も一般的な原因は、染色体の異常によるものであり、特にトリソミー21のような自己染色体異数性が、高齢の母親において特に関連しています(ソース)。
異数性が細胞の振る舞いや機能に与える影響に関する研究も、重要な進展を遂げています。異数性は、細胞の遺伝子コピー数の不均衡を引き起こし、細胞の生理的な状態や振る舞いに深刻な影響を及ぼすことが示されています。異数性の細胞は、通常の細胞とは異なる方法で成長し、分裂し、そして死ぬことがあり、これらの違いが健康や病気の状態にどのように影響するかを理解することは、疾患の治療や管理に新たなアプローチを提供する可能性があります(ソース)。
さらに、染色体の不安定性(CIN)と異数性は、がんにおける人間の免疫系との複雑な相互作用に影響を及ぼします。これらの遺伝的特徴は、がん細胞が免疫系を回避する方法を進化させる一方で、免疫系ががん細胞を認識し攻撃する能力にも影響を与えます。特に、異数性があるがん細胞は、免疫系による監視から逃れたり、免疫系をがんを促進する方向に変える能力を持っています。このような相互作用の理解は、異数性を持つ腫瘍に対する免疫療法の成功に不可欠です(ソース)。
これらの発見は、異数性の研究が人間の健康と病気の理解においてますます重要な役割を果たしていることを示しています。将来的には、これらの知見が新しい治療法や診断法の開発につながることが期待されます。
よくある質問と答え
Q1: 異数性とは何ですか?
A1: 異数性(アネプロイディ)とは、正常な染色体数と異なる数の染色体を持つ状態を指します。人間の場合、通常は46本の染色体(23対)を持ちますが、異数性の状態では、この数が増えたり減ったりしています。
Q2: 異数性の原因は何ですか?
A2: 異数性は、主に細胞分裂の過程で染色体が正しく分配されないことによって生じます。これは、減数分裂または有糸分裂の際に起こり得るエラーです。高齢の母親から生まれた子供によく見られることがあり、その他にも遺伝的要因や外的要因が影響することがあります。
Q3: 異数性がもたらす影響は何ですか?
A3: 異数性は、発達遅延、知的障害、身体的特徴の変化、繁殖能力の低下など、さまざまな影響を及ぼすことがあります。例えば、ダウン症候群(トリソミー21)は、21番染色体が3本存在することによる異数性の一例です。
Q4: 異数性は遺伝しますか?
A4: 異数性が遺伝するかどうかは、その原因やタイプによります。一部の異数性疾患は親から子へ遺伝することがありますが、多くの異数性は生殖細胞の形成過程や初期胚発育の際の偶発的なエラーによって生じるため、必ずしも遺伝するわけではありません。
Q5: 異数性の診断方法は?
A5: 異数性は、染色体分析(カリオタイピング)、フルオレセンス・イン・シチュ・ハイブリダイゼーション(FISH)、配列ベースの手法(例:次世代シーケンシングを含む)など、さまざまな遺伝的検査によって診断されます。妊娠中にはNIPTや羊水検査、絨毛検査を通じて診断されることもあります。
Q6: 異数性の治療法はありますか?
A6: 異数性自体を「治療」する方法はありませんが、異数性によって引き起こされる症状や状態の管理やサポートが可能です。これには、教育的支援、物理療法、言語療法などが含まれます。重要なのは、個々のニーズに応じた適切な介入とサポートを提供することです。
異数性についてのこれらの質問と答えは、基本的な理解を提供するものであり、異数性に関連する状態や症状については、医療専門家との相談が推奨されます。







