目次 [∧]
メッセンジャーRNAとは?
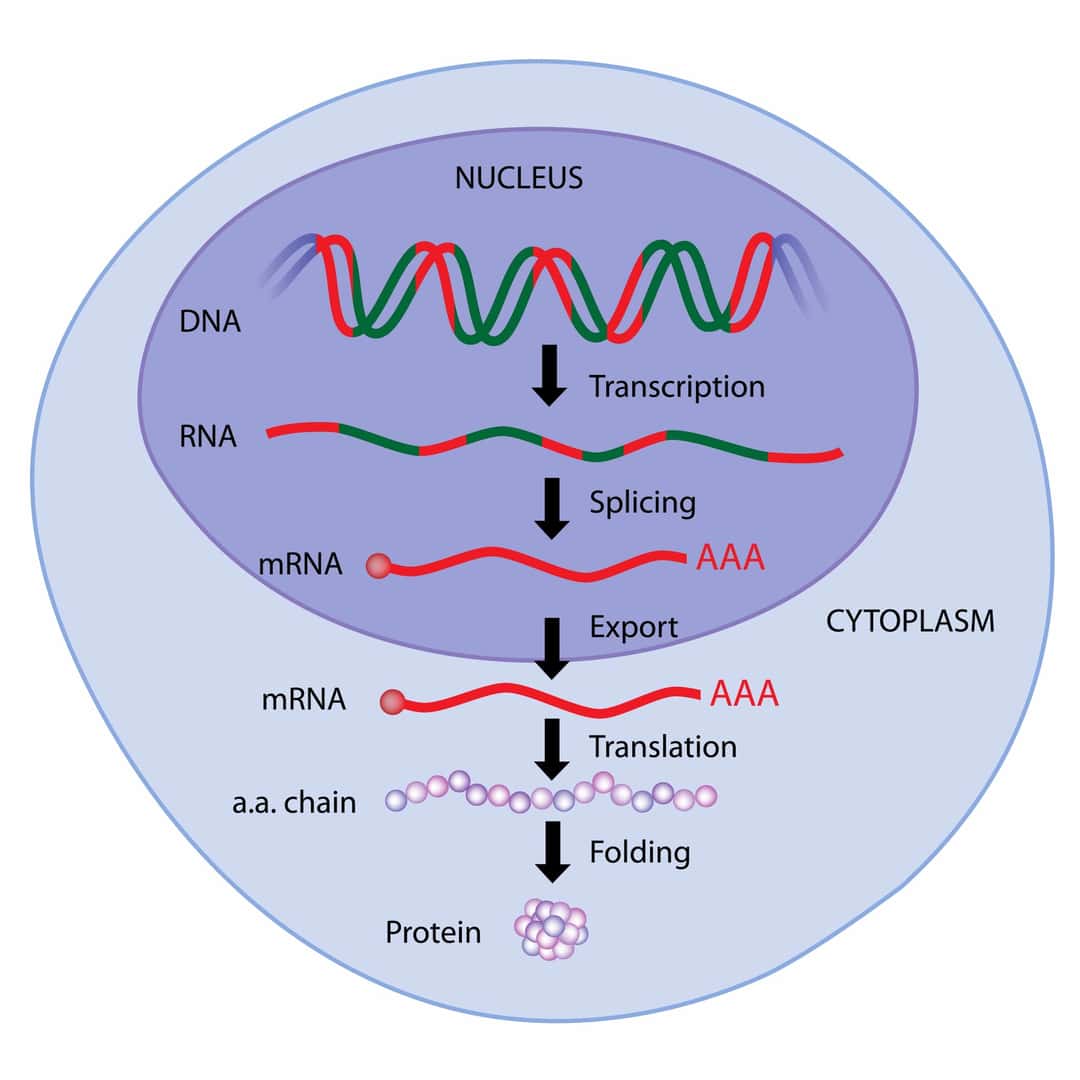
メッセンジャーRNA(mRNA)は、遺伝子の情報をタンパク質の合成場であるリボソームに伝える役割を持つRNAです。このRNAは、DNAの遺伝情報を基にして、RNAポリメラーゼという酵素によって転写されます。
mRNAは、DNAの塩基配列に対応する塩基配列を持ち、この情報をもとにリボソームでタンパク質が合成されます。具体的には、mRNAの塩基配列がアミノ酸の配列に翻訳され、それがタンパク質を形成します。
mRNAの生成過程では、DNAがRNAポリメラーゼによって転写され、その結果としてmRNAが合成されます。この過程で、真核生物ではプレmRNAが生成され、これがさらに加工(スプライシング、キャッピング、ポリアデニル化)を受けて成熟したmRNAとなります。
mRNAは、その構造が不安定で寿命が短いため、遺伝子の発現を迅速に調節することが可能です。また、mRNAベースのワクチンや治療法では、このmRNAを利用して特定のタンパク質を細胞内で合成させ、免疫応答を誘導することがあります。
メッセンジャーRNA(mRNA)の役割とは?
メッセンジャーRNA(mRNA)の役割は、細胞の核内でDNAの持つ遺伝情報を転写し、タンパク質合成の場であるリボソームへとその情報を伝えることです。具体的には、DNAに保持されている遺伝情報がRNAポリメラーゼによってmRNAに転写され、このmRNAが細胞質へと移動してリボソームでタンパク質の合成に利用されます。
mRNAは、タンパク質を作る設計図として機能し、リボソームでの翻訳過程において、その塩基配列に基づいて特定のアミノ酸配列を持つタンパク質が合成されます。このプロセスは、細胞の様々な機能を担うタンパク質を生産するために不可欠です。
また、mRNAはワクチンの分野でも重要な役割を果たしています。mRNAワクチンは、病原体の一部分を作る設計図となるmRNAを体内に導入し、細胞がその設計図に従って病原体のタンパク質の一部を合成します。この合成されたタンパク質は免疫系によって異物と認識され、抗体の産生やT細胞の活性化を通じて免疫応答を引き起こし、将来の感染に備えます。この技術は、新型コロナウイルス感染症(COVID-19)の予防を目的としたワクチンの開発において、初めて公的に承認されたmRNAワクチンで利用されました。
どうしてメッセンジャーRNAが必要なのですか
メッセンジャーRNA(mRNA)は、細胞内でタンパク質を合成するために不可欠な分子です。具体的には、以下のような重要な役割を担っています:
1. 遺伝情報の伝達: DNAは細胞の核に存在し、その情報は直接利用されません。代わりに、DNAの遺伝情報は転写過程を通じてmRNAにコピーされます。このmRNAが核から細胞質へと運ばれ、リボソームで読み取られることによって、遺伝情報がタンパク質という実際の形で表現されます。
2. タンパク質合成の指示: mRNAはリボソームによって読み取られ、その塩基配列に従ってアミノ酸が組み合わされ、タンパク質が合成されます。mRNAの各コドン(3つのヌクレオチドからなる単位)は特定のアミノ酸を指定し、これにより正確なタンパク質が生成されます。
3. 遺伝的制御の調節: 細胞はさまざまなタイプのmRNAを異なる量で生産することによって、必要なタンパク質の種類と量を制御します。これにより、細胞の成長、分裂、応答など、細胞の機能を正確に調節できます。
4. 遺伝的多様性の確保: mRNAは転写後にさまざまな加工を受けることがあります(例:スプライシング)。これにより、一つの遺伝子から異なるタンパク質が複数生成されることが可能となり、遺伝的な情報を効率的に利用することができます。
5. 細胞間のコミュニケーション: 特定のmRNAは細胞外へ放出され、他の細胞に運ばれることがあります。このプロセスを通じて、細胞間で情報を共有し、体全体の調和を取ることが可能です。
これらの機能により、mRNAは生命活動の基本的なプロセスであるタンパク質合成を効果的に支援し、細胞や生物全体の生理的機能の実行に不可欠な役割を果たしています。
メッセンジャーRNA(mRNA)の合成場所
メッセンジャーRNA(mRNA)の合成場所は、生物の種類によって異なります。
1. 原核生物:
– 原核生物では、核を持たないため、mRNAの合成(転写)は細胞質で行われます。
2. 真核生物:
– 真核生物では、mRNAの合成は細胞核内で行われます。具体的には、DNAからの情報がRNAポリメラーゼによってmRNAに転写され、その後、成熟したmRNAは核膜孔を通って細胞質に輸送されます。
このように、mRNAの合成場所は、原核生物と真核生物で異なり、それぞれの細胞の構造に適応した場所で転写が行われます。
mRNAの構造
mRNAには、タンパク質をコードする部分があり、これは翻訳開始コドンであるAUGから始まり、終止コドン(UAA、UGA、UAGの3種類)で終わります。この部分の前には5’末端非翻訳領域(5’UTR)、後ろには3’末端非翻訳領域(3’UTR)があります。真核生物のmRNAでは、5’末端にはキャップ構造があり、3’末端にはポリ(A)鎖が付加されています。これらの構造は、mRNAの安定性やタンパク質の合成に重要な役割を果たします。
ほとんどのmRNA分子の5’末端には、メチル化されたグアニンヌクレオチドが逆向きに付加されています。この構造を「7-メチル-グアノシンの5’キャップ構造(m7GpppX、Xは任意のヌクレオチド)」と呼びます。このキャップ構造は、翻訳の開始因子であるeIF4Eによって認識され、それが他の開始因子をmRNAに引き寄せる役割を果たします。このキャップ構造は、mRNAの翻訳、スプライシング、安定性の向上、さらにはRNAの細胞核からの搬出を促進するなど、細胞内で多くの重要なプロセスに関与しています。eIF4Eは、eIF4F複合体の一部として、または遊離型として存在します。eIF4F複合体には、ATPアーゼ活性とRNAヘリカーゼ活性を持つeIF4Aと、足場タンパク質であるeIF4Gも含まれています。
mRNAの3’末端には、ポリ(A)鎖と呼ばれる200-250ヌクレオチドの長いアデニンの連続があります。このポリ(A)鎖にはポリ(A)鎖結合タンパク質(PABP)が結合します。PABPは、mRNAのポリ(A)テールに直接結合するeIF4G(ユーカリオート起動因子4複合体)の足場として機能するRNA結合タンパク質です。PABPの役割は、mRNAの尾部を分解から守り、mRNAの安定性と産生を促進することです。PABPとeIF4Gが連携しない場合、ポリ(A)テールが付加されず、RNAは迅速に分解される可能性があります。このシステムは、mRNAの寿命と効率的な翻訳を確保するために重要です。
PABP(ポリ(A)鎖結合タンパク質)とeIF4Eは、翻訳開始因子であるeIF4Gを介して複合体を形成し、この結果、mRNAは環状構造を取るようになります。この環状構造は、mRNAの3’非翻訳領域がポリ(A)鎖より近位側でたわんで外れることによってさらに調整されます。このプロセスには翻訳終結因子eRF3も関与し、PABPとeIF4Gとの複合体形成をサポートします。
mRNAがこのように環状化することにより、翻訳終結部位(3’末端)と翻訳開始部位(5’末端)が物理的に近づけられます。この配置は、翻訳を終えたリボソームが終止コドンから次の翻訳開始コドンへ効率的に移行することを容易にし、翻訳の効率を向上させる重要な役割を果たしています。このメカニズムにより、リボソームは迅速に再利用され、タンパク質の合成が効率的に行われるのです。
mRNAの5’末端にあるキャップ構造と3’末端のポリ(A)鎖は、mRNAの安定性を高める重要な役割を果たします。これらの構造は、エキソヌクレアーゼと呼ばれる酵素によるmRNAの分解を阻害します。エキソヌクレアーゼは、RNA分子の端から酵素反応を行ってRNAを分解する酵素ですが、キャップ構造とポリ(A)鎖がこれを妨げることで、mRNA分子はより長く細胞内で機能し続けることができます。これにより、mRNAからのタンパク質合成が効率的に行われることを保証し、細胞の機能維持に寄与しています。
mRNAの翻訳
翻訳は、転写によって生成されたmRNAの塩基配列を読み取り、その指示に従ってアミノ酸を組み合わせてタンパク質を合成するプロセスです。この過程はリボソームという細胞の機構が行います。mRNAのコドン(3つのヌクレオチドから成る単位)がリボソームによって一つずつ読まれ、それぞれのコドンに対応するアミノ酸が運ばれてきます。アミノ酸はペプチド結合で連結され、最終的に特定のタンパク質が合成されます。
真核生物の翻訳プロセスには、開始、伸長、終結の三つの主要な段階が含まれています。以下に各段階の概要を説明します:
● 翻訳開始
翻訳の開始は、メッセンジャーRNA(mRNA)の5’キャップ構造が翻訳の開始因子であるeIF4Eによって認識されることから始まります。この認識は、mRNAとリボソームの小サブユニットとの結合を促進し、翻訳の準備を整える重要な初期ステップです。eIF4Eは、eIF4GとeIF4AとともにeIF4F複合体を形成し、リボソームの小サブユニットがmRNAの5’末端からスキャンを開始し、翻訳の開始コドン(通常はAUG)を見つけるのを助けます。リボソームが開始コドンを認識すると、大サブユニットが結合し、翻訳が開始されます。
● 翻訳伸長
開始コドンで翻訳が開始されると、リボソームはmRNAを沿って移動し、各コドンを読み取って対応するアミノ酸を結合させ、成長中のポリペプチド鎖を伸ばしていきます。これは伸長フェーズと呼ばれ、リボソームはペプチジル転移反応を繰り返しながら、一つのアミノ酸を次々とポリペプチド鎖に追加していきます。伸長過程では、複数のエロンゲーション因子がリボソームの効率的な機能をサポートします。
● 翻訳終結
翻訳終結の過程は、特定の翻訳終結因子によって制御されます。主要な因子には、Eukaryotic translation termination factor 1 (eRF1) と Eukaryotic translation termination factor 3 (eRF3) が含まれます。翻訳終結は以下のステップで進行します:
1. 終止コドンの認識: 終止コドンUAA、UAG、UGAは、eRF1によって認識されます。これらのコドンはリボソームのA部位(アミノアシルtRNAの入口)に位置するときにeRF1によって直接認識されます。
2. ペプチド鎖の解離: eRF1が終止コドンに結合すると、リボソーム内で合成されたペプチド鎖を解離させます。これにより、ポリペプチドはリボソームから放出され、その後の折り畳みや修飾が行われる準備が整います。
3. eRF3の役割: eRF3はGTP結合タンパク質であり、GTP結合型のeRF3はeRF1と複合体を形成してリボソームに結合します。この複合体は、GTPの加水分解が促進されることによって活性化され、加水分解が完了するとGDP型に構造が変化し、これがeRF1からの分子スイッチとしての解離を促します。
このように、eRF1とeRF3は連携してリボソーム上での翻訳プロセスの終結を効率的に行い、タンパク質の合成を正確に終了させる重要な役割を果たしています。このプロセスを通じて、正確なタンパク質が合成され、細胞の機能と生存に必要なタンパク質が供給されます。
これらの過程を通じて、mRNAの遺伝情報は具体的なタンパク質として翻訳され、細胞の構造や機能を維持するために使用されます。

この記事の筆者:仲田洋美(医師)




