目次

📚 入門シリーズ
この記事は「遺伝病を理解するためのヒトゲノム入門編」の1本です。全記事の一覧と学ぶ順番はヒトゲノム入門編・索引ページからご覧いただけます。
📍 クイックナビゲーション
「ゲノム病(genomic disorder)」とは、染色体のごく一部が抜け落ちたり(微細欠失)、余分に増えたり(微細重複)することで起こる病気の総称です。原因は1文字だけの遺伝子変異ではなく、数十万〜数百万塩基という「かたまり」単位のコピー数の増減にあります。なぜ特定の場所で繰り返し起こるのか、どんな症候群があるのか、そしてマイクロアレイ(CMA)やNIPTでどこまで分かるのかを、遺伝専門医の視点から一般の方にもわかるように整理します。
Q. ゲノム病とは何ですか?まず結論だけ知りたいです
A. 染色体の一部が「かたまり」で欠けたり(微細欠失)増えたり(微細重複)することで、その中にある用量に敏感な遺伝子の量が過不足になり、発達や臓器形成に影響が出る病気のグループです。その多くは、ヒトゲノムに散らばるよく似た反復配列(LCR)のあいだで起こる組換えミスによって、決まった場所で繰り返し発生します。診断は染色体マイクロアレイ(CMA)が中心で、22q11.2欠失症候群などは出生前にNIPTでもスクリーニングできるようになってきました。
- ➤正体はCNV → 原因は1塩基の変異ではなく、数十万〜数百万塩基のコピー数の増減(欠失・重複)
- ➤起こりやすい場所がある → LCR(低コピー反復配列)にはさまれた領域で、同じ欠失・重複が繰り返し生じる
- ➤欠失は重く、重複は軽い傾向 → 遺伝子が半分になる「ハプロ不全」の影響が大きい
- ➤代表例 → 22q11.2欠失(ディジョージ)、ウィリアムズ、プラダー・ウィリ/アンジェルマンなど
- ➤診断はCMAが中心 → 発達の遅れなどでは核型検査(約3%)よりCMA(15〜20%)の診断率が高い
1. ゲノム病とは何か:CNV(コピー数変異)が主役の病気
遺伝性の病気というと、多くの方は「DNAの1文字が書き換わる」タイプの変異(点変異)を思い浮かべるかもしれません。ハンチントン病や嚢胞性線維症のように、1つの遺伝子の中の小さな変化で起こる「メンデル遺伝病」がその典型です。これに対してゲノム病(genomic disorder)は、染色体上のまとまった長さのDNAそのものが、丸ごと失われたり(欠失)、余分に増えたり(重複)することで起こります[1]。この「かたまり」の大きさは、数千塩基(kb)から数百万塩基(Mb)にまで及びます。
このような、ゲノムの一部の量が増えたり減ったりする変化を、まとめてコピー数変異(CNV:Copy-Number Variation)と呼びます。ゲノム病の本体は、この不均衡なCNV——つまり、本来2本あるべき遺伝子が1本になる「微細欠失」や、3本に増える「微細重複」です。顕微鏡で見える大きな染色体異常(ダウン症候群のような数の異常)とは異なり、ゲノム病の多くは光学顕微鏡では見えないほど小さく、専門的な検査が必要になります。
💡 用語解説:CNV(コピー数変異)
ヒトの染色体は、父由来・母由来の2本1組になっています。ふつうは、その中の各領域が「2コピー」ずつ存在します。CNVとは、この本来2コピーであるべきDNAの一部が、1コピー(欠失)や3コピー(重複)になってしまう変化のことです。健康に影響しない「ただの個人差」としてのCNVも数多くありますが、用量に敏感な遺伝子を含む領域でCNVが起きると、病気の原因になります。ゲノム病は「病気を起こすCNV」によって生じる疾患群だといえます。
ゲノム病という概念が確立したのは比較的最近のことです。ヒトゲノム全体を調べる技術が発達し、「病気は、ゲノムの構造そのもの(アーキテクチャ)に由来する再構成によっても起こる」ことが分かってきました[1]。つまりゲノム病は、DNAの「文字」の間違いではなく、DNAの「並び方・構造」に起因する病気だと位置づけられます。この考え方は、後で説明する診断の進歩(染色体マイクロアレイの登場)と深く結びついています。
2. なぜ決まった場所で起こる?ゲノム構造とLCR
ゲノム病のいちばん重要な特徴は、同じ場所で、同じサイズの欠失・重複が、無関係な多くの患者さんで繰り返し起こるという点です。これは偶然ではありません。ヒトゲノムには、配列が非常によく似た(同一性が高い)DNAのブロックが多数散らばっており、これが「間違い」を誘発する土台になっているのです。この高度に重複したブロックを低コピー反復配列(LCR:Low-Copy Repeats)、あるいは分節重複(SD:Segmental Duplications)と呼びます[4]。
💡 用語解説:LCR(低コピー反復配列)・分節重複
LCRは、長さがおおむね10〜400キロ塩基(kb)、配列の同一性が95〜97%以上という、互いによく似たDNAのかたまりです[4]。ヒトゲノム全体の約5〜6.6%を占めるとされ、進化の過程で新しい遺伝子を生み出す原動力になった一方で、構造的に不安定な「弱点」にもなっています。あまりに似ているために、細胞が「これは同じ場所だ」と勘違いして貼り合わせてしまい、間の領域が欠けたり増えたりするのです。AluやLINEといった、より短くゲノム全体に散らばる古典的な反復配列とは区別されます。
重要なのは、LCRの「向き」と「配置」です。LCRが同じ向きに並んでいるか、逆向きに並んでいるか、あるいは同じ染色体上か別の染色体上かによって、生じる再構成(欠失・重複・逆位)の種類が決まります[3]。次の章で、この物理的なしくみを具体的に見ていきます。
3. 発症のしくみ:NAHR・末端結合・複製エラー
再発性の欠失・重複を作る「NAHR」
同じ場所で繰り返し起こる(再発性の)欠失・重複を生み出す主役が、非対立遺伝子間相同組換え(NAHR:Non-Allelic Homologous Recombination)です[3]。精子や卵子がつくられる減数分裂のとき、父由来と母由来の染色体は正しいパートナー同士で並ぶはずですが、LCRがあまりに似ているために、本来のペアではない場所(非対立の場所)でずれて貼り合わさることがあります。このズレた状態で組換えが起きると、間の領域が失われたり増えたりします。
💡 用語解説:NAHRとは(LCRの向きで結果が変わる)
NAHRは、よく似たLCR同士が誤ってペアになって組換えを起こす現象です。LCRの向きによって、生じる結果が次のように変わります[3]。
- ▸同じ向き(direct)のLCR間:間の領域が一方から失われ他方で増え、欠失と重複が同時に生じます。
- ▸逆向き(inverted)のLCR間:間の領域の向きだけが反転する逆位が生じます(多くはコピー数は変わりません)。
LCRの向きと、生じる再構成のちがい
同じ向きのLCR
— 領域 —
LCR ▶
↓
欠失(1コピーに)+
重複(3コピーに)
逆向きのLCR
— 領域 —
◀ LCR
↓
逆位(向きが反転/
多くはコピー数不変)
LCRの配置(向き)が、欠失・重複か、逆位かを決める。逆位はそれ自体は無症状のことも多いが、次の世代でさらなる欠失・重複を起こしやすくする「素因」になることがある。
この「相反的な欠失・重複」を最初に示した古典例が、末梢神経の病気であるシャルコー・マリー・トゥース病1A型(CMT1A)と、その相反の欠失で起こる遺伝性圧脆弱性ニューロパチー(HNPP)です[3][9]。どちらも17番染色体短腕(17p12)にあるPMP22という同じ遺伝子が関わり、その領域が重複するとCMT1A、欠失するとHNPPになります。同じ場所の「増える/減る」が、鏡に映したように異なる病気を生む——この関係が、ゲノム病を理解する原型になりました。
患者さんごとに切れ目が違う「非再発性」のしくみ
一方で、患者さんごとにサイズや境界がバラバラな「非再発性」の再構成もあります。これらは、DNAが切れたときの修復ミスによって生じます。ひとつは非相同末端結合(NHEJ)で、二本鎖切断が起きた末端を、相同性を頼らずに直接つなぎ直すしくみです。もうひとつはマイクロホモロジー媒介末端結合(MMEJ)で、ごく短い相補配列(マイクロホモロジー)を手がかりに結合します。非再発性の欠失の切断点を詳しく調べた研究では、接合部の91.7%にマイクロホモロジーが確認されており、こうした微小相同性を利用した修復が主導していることが分かっています[7]。
さらに複雑な再構成(欠失と重複の混在や、部分的な三重複など)を説明するのが、DNA複製のエラーに基づくFoSTeS/MMBIRというモデルです[5][6]。複製の途中でフォーク(複製の進行点)が止まると、合成途中の鎖が別の場所へ「乗り換えて」複製を再開することがあり、その結果、離れた領域どうしが強引につなぎ合わされて複雑なCNVが生じます。連続的な複製の滑り(SRS/BISRS)も、非再発性の微細欠失の形成に関わることが確認されています[7]。私たちの生命維持に不可欠な複製・修復のしくみそのものが、同時にゲノムを不安定にする「両刃の剣」でもある、というわけです。
4. 欠失と重複の「非対称性」:なぜ欠失のほうが重いのか
相反の欠失症候群と重複症候群を比べると、臨床上の大きな「非対称性」が見えてきます。一般に、欠失症候群のほうが、対応する重複症候群よりも発症頻度が高く、症状も重い傾向があります[2]。これは、必須のタンパク質が「50%に減る(ハプロ不全)」ことは体にとって致命的になりやすいのに対し、「150%に増える」ことは、細胞内の分解・緩衝のしくみである程度受け止められやすいことを反映しています。
💡 用語解説:ハプロ不全(用量感受性)
ハプロ不全とは、2つあるべき遺伝子のうち1つが失われたことで、つくられるタンパク質が半分になり、それだけで機能が足りなくなって症状が出る状態です。すべての遺伝子がこうなるわけではなく、「量に敏感な(用量感受性の)遺伝子」で起こります。ゲノム病では、欠失した領域の中にこうした用量感受性遺伝子が含まれているかどうかが、症状の重さを大きく左右します。
この非対称性のなかでも、欠失と重複が「鏡像(相反)」の表現型をとる例として有名なのが、17番染色体短腕(17p11.2)の約3.7Mb領域で起こるスミス・マギニス症候群(SMS:欠失)とポトキ・ルプスキ症候群(PTLS:重複)です[8]。SMSでは幼児期以降に過食・肥満をきたしやすいのに対し、PTLSでは乳児期からの筋緊張低下と摂食不良でやせ型になりやすい、という真逆のエネルギー代謝を示します。この鏡像の大部分を単独で支配しているのが、領域内のRAI1という転写因子です。RAI1が1コピー(ハプロ不全)か3コピー(過剰)かの差が、全身の表現型を反転させる引き金になっていることが、マウスモデルでも示されています[8][9]。
5. 代表的な微細欠失・重複症候群
ゲノム病には数十の症候群が知られています。表現型(症状)は、欠失・重複した領域にどの用量感受性遺伝子が含まれるかで決まります。ここでは代表的なものを整理します。多くは新生突然変異(de novo変異)——つまり両親にはなく、お子さんで初めて生じる変化——として起こります。
このなかで、ゲノム病の一般原則をよく示すのが22番染色体長腕(22q11.2)です。次の図は、分節重複(LCR)間の組換えによって、この領域で欠失・重複・再構成が起こるしくみを示しています。ディジョージ症候群では、一方の染色体の約3Mbの領域が失われ、その中の数十の遺伝子(TBX1など)のハプロ不全が症状につながります。
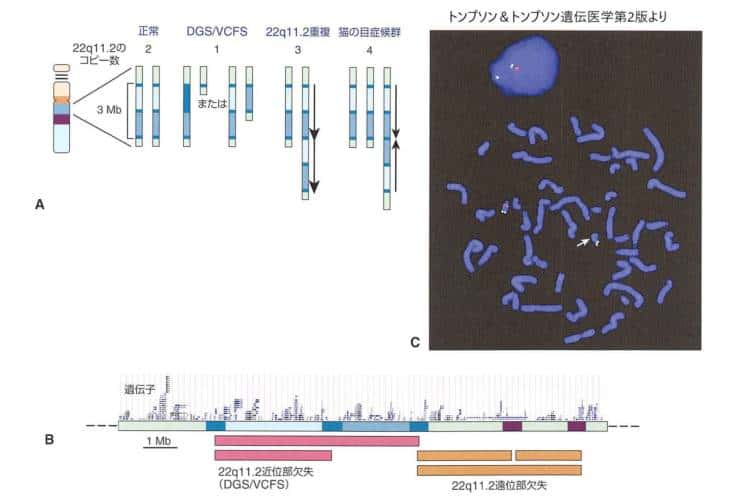
分節重複(LCR)間の組換えにより、22q11.2で欠失・重複・逆位が生じるしくみ。正常核型は2コピーの領域をもち、それぞれの中に多数の分節重複を含む。ディジョージ症候群/口蓋帆・心・顔症候群では約3Mbが欠失し、多くの遺伝子が失われる。
22q11.2欠失症候群のTBX1遺伝子の欠失は先天性心疾患、とくに左室流出路の形態異常と関連が深いと考えられています。一方、22q11.2の重複ははるかにまれで、特徴的な形態異常や先天性疾患を示す22q11.2重複症候群となります。この22q11.2の原則は、他の多くのゲノム病にも共通します。こうした複数の隣接する遺伝子がまとめて失われて起こる病気を隣接遺伝子症候群とも呼びます。
6. インプリンティング型のゲノム病:PWSとAS
🔍 関連記事:ゲノムインプリンティング関連疾患/片親性ダイソミー/アンジェルマン症候群
15番染色体長腕(15q11-q13)は、「どちらの親から受け継いだか」で遺伝子の働きが変わるゲノムインプリンティング(遺伝子刷り込み)の集積地です。この領域の遺伝子は、片方の親由来のコピーだけが働くように調整されているため、生理的に「片方だけが活きている」状態になっています[10]。そのため、同じ領域が失われても、失われたのが父由来か母由来かで、まったく別の病気になります。
💡 用語解説:ゲノムインプリンティング
ふつうの遺伝子は父由来・母由来の両方が働きますが、一部の遺伝子はDNAメチル化という「目印」によって、片方の親由来のコピーだけが働くように制御されています。これがインプリンティングです。どちらの親から来たかで機能が違うため、欠失や片親性ダイソミー(両方のコピーが同じ親由来になること)が起きると、同じ場所でも異なる病気が生じます。
プラダー・ウィリ症候群(PWS)は、父由来の15q11-q13が失われる(または母親由来の片親性ダイソミーが起こる)ことで発症します。この場合、通常は父由来からのみ働くSNORD116などの遺伝子群の発現がゼロになり、乳児期の強い筋緊張低下・哺乳障害から、幼児期以降の制御しにくい過食・肥満へと進む2相性の経過をとります[10]。一方、アンジェルマン症候群(AS)は、母由来の同じ領域が失われることで発症し、重度の知的障害、実用的な言葉の欠如、失調性の動き、理由のない笑いなどを特徴とします。ASでは、脳の神経細胞で母由来のみが働くUBE3A遺伝子が失われることが本質です[10]。
📌 ポイント:インプリンティング型(PWS/AS)が疑われるときの第一選択検査は、コピー数を見るマイクロアレイではなく「メチル化解析」です。欠失・片親性ダイソミー・刷り込み異常のいずれでも共通して検出できるためです[10]。

7. 診断の進歩:核型 → FISH → マイクロアレイ → NGS
ゲノム病を見つける検査は、解像度と網羅性の両面で大きく進化してきました。従来の標準核型分析(G分染法)は、大きな染色体異常(おおむね5〜10Mb以上)を見るには有用ですが、ゲノム病の主戦場である1〜3Mb以下の微細な変化を捉えるには解像度が足りません。特定の場所を狙い撃ちで確認するFISHも、ゲノム全体を見渡すことはできません。
これを一変させたのが染色体マイクロアレイ(CMA)です。ゲノム全体のコピー数の増減を、数kbレベルの高い解像度で網羅的にスキャンできます。国際的なガイドラインは、原因不明の発達の遅れ・知的障害、自閉スペクトラム症、多発奇形をもつお子さんに対して、CMAを第一選択(ファーストライン)の検査とすることを強く推奨しています[11]。その根拠は診断率の高さにあり、核型分析の診断率が約3%(既知の大きな異常を除く)にとどまるのに対し、CMAの診断率は15〜20%に達します[11]。実際の臨床研究でも、CMAで異常が見つかった割合は約20%で、そのうち最も多い所見は16p11.2の微細欠失であったと報告され、専門医紹介や心臓・腎臓の精査など、その後の医療方針の変更に直結したことが示されています[12]。
💡 用語解説:染色体マイクロアレイ(CMA)
CMAは、ガラス基板上にびっしり並べた無数のDNAプローブに、患者さんのDNAを反応させて、ゲノム全体のコピー数の過不足を一度に読み取る検査です。顕微鏡では見えない微細な欠失・重複を高解像度で検出できます。ただし、コピー数の増減を伴わない均衡型の転座や逆位、ごく低い割合のモザイク、1文字レベルの点変異は検出できません。こうした場合には核型分析や次のNGSが必要になります。詳しくはCMA検査結果の見方もご覧ください。
診断の最前線は、さらに次世代シーケンサーによる全エクソーム解析(WES)・全ゲノム解析(WGS)へと移りつつあります。発達の遅れ・知的障害を対象に102の研究(合計55,752人)をまとめたメタ解析では、WESの診断率は37%(95%信頼区間33〜41%)で、CMAの19%(16〜21%)を大きく上回りました[14]。これは、CMAでは捉えられない「1つの遺伝子の点変異」まで同じ検査で拾えるためで、専門家の合意声明はWESを神経発達症の第一選択検査に位置づけています[15]。さらに、低カバレッジ全ゲノム(low-pass WGS/CNV-seq)は、CMAが検出したCNVをすべて検出しつつ、追加のCNVも同定できることが前向き研究で示されており[16]、CNV検出と点変異の解析を1つに統合した検査への流れが進んでいます。
なお、ゲノム病の治療の多くは対症療法や合併症の管理が中心ですが、根本原因に介入する研究も進んでいます。CRISPR-Cas9などのゲノム編集や遺伝子治療は一部の単一遺伝子病で臨床応用が始まっていますが、大きな領域の欠失・重複を対象とするゲノム病では、現時点では基礎・研究段階にとどまります。
8. 出生前スクリーニング:NIPTと22q11.2欠失
母体の血液に含まれる、胎盤由来のセルフリーDNA(cfDNA)を解析するNIPT(非侵襲的出生前スクリーニング)は、21・18・13トリソミーなどの数の異常を超えて、微細欠失、とくに22q11.2欠失症候群の検出へと広がっています。その臨床的な有用性を検証した大規模研究が、世界21施設・2万人以上の妊婦を対象とした前向き研究「SMART試験」です[13]。
💡 用語解説:cfDNAと陽性的中率(PPV)
cfDNAは、妊婦さんの血液中を流れる、胎盤由来の細かく断片化したDNAです。採血だけで胎児の染色体の状態をスクリーニングできます。陽性的中率(PPV)とは、「検査で陽性となった人のうち、実際に本当に該当していた人の割合」です。PPVが53%なら、陽性の約2人に1人が真の陽性という意味になります。スクリーニング検査である以上、陽性の場合は羊水検査などの確定検査が必要です。
SMART試験が示した22q11.2欠失スクリーニングの主な数値
SNP型cfDNA(Panorama)を用いた前向き検証[13]
1/1,524
検出頻度(この集団)
83.3%
感度
99.9%超
特異度
53%
陽性的中率(PPV)
最も一般的な2.5〜3Mbの欠失はすべて検出し、22q11.2欠失全体の83%を検出。従来の母体血清マーカーによるダウン症検査のPPV(約3%)を大きく上回る[13]。
この結果には重要な意味があります。SMART試験では、出生前に22q11.2欠失が確認された胎児のうち、妊娠初期の超音波検査では異常が全く見られなかったことが示されました[13]。つまり、超音波だけでは早期に気づけない微細欠失を、cfDNAが妊娠早期に把握できる可能性があるということです。出生前にこの欠失が分かっていれば、重い心奇形への出生直後の対応、免疫不全を想定した生ワクチンの取り扱い、低カルシウム血症への備えなど、出生後の医療体制をあらかじめ整えることにつながります。一方で、22q11.2以外のより稀な微細欠失では現時点でPPVが低めであることも報告されており、検査の限界と併せた丁寧な説明が欠かせません[13]。
📌 親世代への波及:22q11.2欠失症候群は常染色体顕性(優性)遺伝の形をとりますが、その多くはお子さんで初めて生じる新生突然変異(de novo変異)です。ただし、症状が軽く未診断のまま成人した保因者の親も一定数存在します。胎児のNIPT陽性をきっかけに、親自身の潜在的な欠失が初めて分かることもあり、家系全体の遺伝カウンセリングにつながります。
9. よくある誤解
誤解①「ゲノム病は遺伝するから親に原因がある」
代表的なゲノム病の多くは、両親にはない新生突然変異(de novo変異)としてお子さんに生じます。親の育て方や生活習慣が原因ではありません。ただし一部に親からの遺伝例もあるため、正確な再発リスクは検査と遺伝カウンセリングで確認します。
誤解②「重複(コピーが増える)なら害はない」
重複は欠失より軽い傾向はありますが、無害とは限りません。遺伝子が過剰に働くことで発達の遅れや症状を起こす重複症候群もあります。「増える」「減る」のどちらも、含まれる遺伝子次第で影響が変わります。
誤解③「ふつうの血液検査や核型でわかる」
ゲノム病の多くは顕微鏡で見える大きさより小さいため、標準核型では捉えられません。マイクロアレイ(CMA)や、疑う病気に応じた専門的な検査が必要です。どの検査を選ぶかが診断の分かれ目になります。
誤解④「PWS/ASもマイクロアレイで確定できる」
インプリンティング型(プラダー・ウィリ/アンジェルマン症候群)は、欠失だけでなく片親性ダイソミーや刷り込み異常でも起こるため、第一選択はメチル化解析です。CMAでは一部のタイプを見逃す可能性があります。
よくある質問(FAQ)
🏥 染色体・ゲノムの検査のご相談
微細欠失・重複症候群やその出生前スクリーニングについて
マイクロアレイ・NIPT・確定検査の選び方まで
臨床遺伝専門医が在籍するミネルバクリニックにお気軽にご相談ください。
参考文献
- [1] Shaw CJ, Lupski JR. Implications of human genome architecture for rearrangement-based disorders: the genomic basis of disease. Hum Mol Genet. 2004. [Oxford Academic]
- [2] Lupski JR, Stankiewicz P. Genomic disorders: molecular mechanisms for rearrangements and conveyed phenotypes. PLoS Genet. 2005. [PLOS Genetics]
- [3] Kumar D. Disorders of the genome architecture: a review. Genomic Med. 2008. [PMC2694859]
- [4] Genome Architecture and Its Roles in Human Copy Number Variation. Genomics Inform. [Genomics & Informatics]
- [5] Zhang F, et al. The DNA replication FoSTeS/MMBIR mechanism can generate genomic, genic and exonic complex rearrangements in humans. Nat Genet. 2009. [PMC4461229]
- [6] Hastings PJ, Ira G, Lupski JR. A microhomology-mediated break-induced replication model for the origin of human copy number variation. PLoS Genet. 2009. [PLOS Genetics]
- [7] Microhomology-Mediated Mechanisms Underlie Non-Recurrent Disease-Causing Microdeletions of the FOXL2 Gene or Its Regulatory Domain. PLoS Genet / PMC3597517. [PMC3597517]
- [8] Neurodevelopmental Disorders Associated with Abnormal Gene Dosage: Smith-Magenis and Potocki-Lupski Syndromes. J Pediatr Genet / PMC4918721. [PMC4918721]
- [9] Nonrecurrent 17p11.2p12 Rearrangement Events that Result in Two Concomitant Genomic Disorders: The PMP22-RAI1 Contiguous Gene Duplication Syndrome. Am J Hum Genet / PMC4667131. [PMC4667131]
- [10] Prader-Willi, Angelman, and 15q11-q13 duplication syndromes. Pediatr Clin North Am / PMC4449422. [PMC4449422]
- [11] Miller DT, et al. Consensus statement: chromosomal microarray is a first-tier clinical diagnostic test for individuals with developmental disabilities or congenital anomalies. Am J Hum Genet. 2010. [PMC2869000]
- [12] Chromosomal Microarray Analysis as a First-Tier Clinical Diagnostic Test… A Prospective Multicenter Study. Ann Lab Med / PMC6340852. [PMC6340852]
- [13] Dar P, et al. Cell-free DNA screening for prenatal detection of 22q11.2 deletion syndrome (SMART study). Am J Obstet Gynecol. 2022. [AJOG]
- [14] Diagnostic and clinical utility of exome sequencing and chromosomal microarray in children with GDD/ID: a meta-analysis. Ann Med. 2025. [PMC12777888]
- [15] Srivastava S, et al. Meta-analysis and multidisciplinary consensus statement: exome sequencing is a first-tier clinical diagnostic test for individuals with neurodevelopmental disorders. Genet Med. 2019. [Genetics in Medicine]
- [16] Wang H, et al. Low-pass genome sequencing versus chromosomal microarray analysis: implementation in prenatal diagnosis. Genet Med. 2020. [PMC7042067]






