目次
- 1 1. コヒーシンの「2つの顔」:接着装置であり、分子モーターでもある
- 2 2. コヒーシンの分子構造:巨大なリングをつくる部品たち
- 3 3. ループ押し出しの仕組み:「スイング&クランプ」というナノモーター
- 4 4. CTCFと転写マシナリー:ループの「境界」はどう決まるのか
- 5 5. アセチル化サイクルとWAPL:コヒーシンの「滞在時間」を決める化学スイッチ
- 6 6. 減数分裂におけるコヒーシン:2段階の分裂を成功させる巧妙な制御
- 7 7. コヒーシノパチー:コヒーシンの異常が起こす先天性疾患と遺伝診療
- 8 8. がんとコヒーシン:STAG2という「がん抑制因子」
- 9 9. よくある誤解
- 10 よくある質問(FAQ)
- 11 参考文献
- 12 関連記事

📍 クイックナビゲーション
細胞の核のなかには、身長よりもはるかに長い約2メートルのDNAが折りたたまれて収まっています。この長大なDNAを、細胞分裂のときには姉妹染色体を束ねて正しく分配し、ふだんは必要な遺伝子だけを読み出せるように立体的に整理している立役者が「コヒーシン」というタンパク質の複合体です。このページでは、コヒーシンの構造とはたらき、減数分裂での特別な動き、そしてコルネリア・デ・ランゲ症候群などのコヒーシン関連疾患やがんとの関わりまでを、遺伝専門医の視点でやさしく解説します。
Q. コヒーシンとは何ですか?まず結論だけ知りたいです
A. コヒーシンは、複製された姉妹染色体どうしを分裂の直前まで束ねておく「接着装置」であり、同時にDNAをループ状に手繰り寄せてゲノムの3D構造をつくる「分子モーター」でもある、リング状のタンパク質複合体です。このはたらきが乱れると、コルネリア・デ・ランゲ症候群などの先天性疾患(コヒーシノパチー)や、膀胱がん・白血病などのがんに関わることが知られています。
- ➤2つの顔 → 「姉妹染色体の接着」という維持の機能と、「DNAループ形成」という制御の機能をあわせ持つ
- ➤構成部品 → SMC1A・SMC3・RAD21・STAG1/STAG2などが巨大なリングを組み立てる
- ➤動く仕組み → 「スイング&クランプ」でDNAを一方向に手繰り寄せ、ループを大きくしていく
- ➤先天性疾患 → NIPBL・SMC1A・RAD21・HDAC8などの変化がコヒーシノパチーを起こす
- ➤がんとの関係 → STAG2は多くのがんで失われる「がん抑制因子」として注目されている
1. コヒーシンの「2つの顔」:接着装置であり、分子モーターでもある
コヒーシン(cohesin)は、進化のうえで酵母からヒトまで高度に保存された、複数の部品からできた大きなタンパク質の複合体です。もともとは、DNAが複製されたあとにできる「姉妹染色体(姉妹染色分体)」どうしを、細胞分裂の後半までしっかり束ねておく「接着装置」として発見されました[1]。この接着があるおかげで、分裂のときに染色体を引っぱる糸(紡錘体)が両側から均等に力をかけられ、染色体が娘細胞に正確に1本ずつ配られます。
💡 用語解説:姉妹染色分体(しまいせんしょくぶんたい)
細胞が分裂する準備としてDNAをコピー(複製)すると、まったく同じ内容の染色体が2本、ぴったり寄り添った状態でできます。この2本を姉妹染色分体と呼びます。分裂のときには、この2本を1本ずつ2つの娘細胞に分け合う必要があります。コヒーシンは、分け合う「その瞬間」まで2本を離れないよう束ねておく糊(のり)の役割を果たしています。
ところが近年、コヒーシンにはもう一つの重要な顔があることがわかってきました。細胞が分裂していない時期(間期)に、コヒーシンはみずからDNAを手繰り寄せて「DNAループ」をつくる分子モーターとしてはたらいているのです[3]。この「ループ押し出し」という動きによって、ゲノムはトポロジー関連ドメイン(TAD)と呼ばれる区画に整理され、遠く離れた場所にある遺伝子のスイッチ(エンハンサー)と遺伝子本体(プロモーター)が正しく出会えるようになります。つまりコヒーシンは、どの遺伝子をいつ読み出すかという遺伝子発現のプログラムそのものを左右しています[4]。
この「染色体を物理的につなぐ維持の機能」と「ゲノムの立体構造を組み替える制御の機能」という2つの顔をあわせ持つからこそ、コヒーシンの部品に変化が起きると、発達の障害からがんまで、実にさまざまな病気につながります。似たリング構造を持つコンデンシンが主に分裂期の染色体を「凝縮」させる役割に特化しているのとは対照的に、コヒーシンは接着と制御の両面を担う多機能プレーヤーだといえます。
💡 用語解説:DNAループとTAD(トポロジー関連ドメイン)
長いDNAを、ところどころで輪(ループ)にして手繰り寄せると、ゲノムはいくつかの「区画」に分かれます。この区画がTADです。TADの内側では遺伝子とそのスイッチが活発にやりとりでき、区画をまたいだ余計な干渉は起きにくくなります。コヒーシンはこのループとTADをつくる主役で、ゲノムを「読みやすいフォルダ」に整理していると考えるとイメージしやすいでしょう。くわしくは染色体の構造(TAD・ループ)のページもご覧ください。
2. コヒーシンの分子構造:巨大なリングをつくる部品たち
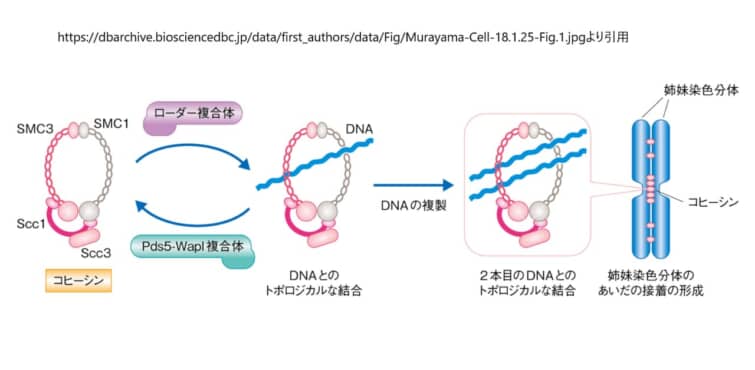
コヒーシンの基本は、DNAを物理的に包み込むことができる巨大なリング(環)状の構造です[2]。このリングは、大きく分けて3種類の部品からできています。骨組みをつくる「SMCタンパク質」、そのすき間を橋渡しして輪を閉じる「クレイシン」、そしてリングのはたらきを調節する「付随タンパク質」です。
骨組みを担うのがSMC1AとSMC3という2本の長いタンパク質です。それぞれが約50ナノメートルにも達する長い腕(コイルドコイル)を持ち、片方の端には輪の「ちょうつがい」にあたるヒンジ、もう片方の端にはエネルギー源であるATPを結合する「ヘッド」があります。SMC1AとSMC3はヒンジどうしでしっかりつながり、全体としてV字型の骨格をつくります。このV字の開いた口を閉じるのが、クレイシンと呼ばれるRAD21です。RAD21の両端がSMC3側とSMC1A側のヘッドに結合することで、3つの部品からなる閉じたリングが完成します。
この基本リングに、STAG1またはSTAG2という部品がRAD21を介して結合します。STAG1とSTAG2は互いにどちらか一方だけが入る関係にあり、染色体のうえで役割分担をしています。STAG2を含むコヒーシンはゲノム全体の大部分を占めて染色体の腕やセントロメア領域の接着を担い、STAG1を含むコヒーシンはテロメア(染色体の末端)の接着維持を得意としています。興味深いことに、STAG2が失われた細胞では、代わりにSTAG1が増えてその穴を埋めようとします。この関係は、のちに述べるがん治療のヒントにもなっています。
リングを載せる係と外す係:ローダーと解離因子
コヒーシンは、ただDNAのそばにあるだけでは働けません。DNAにリングを「載せる」専用の係と、いったん載ったリングを「外す」係がいて、両者のバランスでコヒーシンの居場所と滞在時間が決まります[3]。載せる係がNIPBL–MAU2という複合体(コヒーシン・ローダー)で、外す係がWAPLとPDS5A/PDS5Bです。このNIPBLこそ、コルネリア・デ・ランゲ症候群でもっとも多く変化が見つかる遺伝子であり、コヒーシンの搭載とループ押し出しの両方を推進する中心人物です。
3. ループ押し出しの仕組み:「スイング&クランプ」というナノモーター
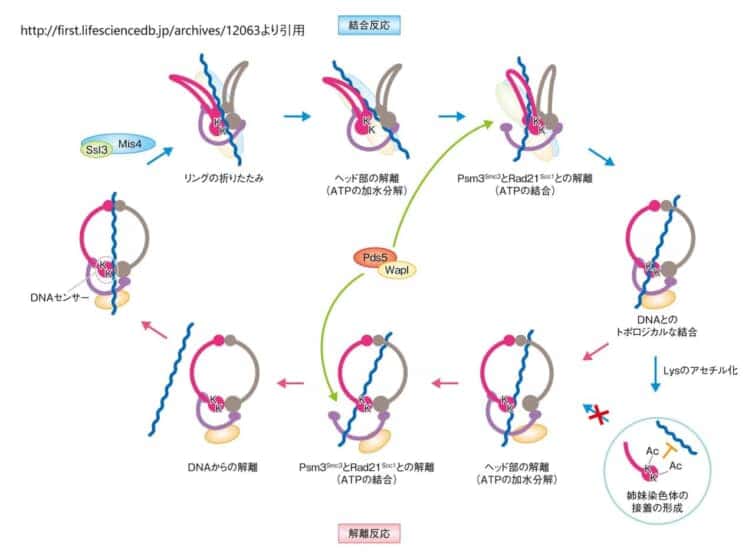
コヒーシンがDNAループをつくる過程は、いくつかの段階に分かれています。まずローダーであるNIPBL–MAU2の助けとATPの力で、コヒーシンは「DNAグリップ状態」という中間の形をとります[7]。このときリングの一部の扉が一時的に開いてDNAが内側に入り込み、SMCのヘッド側とヒンジ側の2つのモジュールがDNAを両側からしっかり挟み込みます。こうして極めて安定した「つかんだ状態」ができあがります。
つかんだあと、コヒーシンは「スイング&クランプ(振って、はさむ)」という動きでDNAの上を進み、ループを少しずつ大きくしていきます[5]。高速で動きを観察できる特殊な顕微鏡(高速原子間力顕微鏡)を使った研究では、コヒーシンの腕が大きく伸び、その先端のヒンジが約50ナノメートル前方へと自発的に「スイング」する様子がとらえられました[6]。スイングによって前方のDNAをつかまえ、ATPの結合と連動してヘッドの上で「クランプ(はさみ込み)」します。ATPが分解されるとヘッドどうしが離れ、はさみ込みが解けて1サイクルが完了します。これを繰り返すことで、コヒーシンはDNAを一方向に手繰り寄せていきます。
ループ押し出しの1サイクル(スイング&クランプ)
①〜④を何度も繰り返すことで、コヒーシンはDNAをたぐり寄せ、ループを連続的に大きくしていきます。
なお、ヘッドの扉を通過してDNAがリングの内側に完全に取り込まれた場合には、姉妹染色体を束ねる「接着」状態になります。一方、扉の通過に失敗した場合には、ヒンジの往復運動が一種のラチェット(逆戻りしない歯車)として働き、リングにDNAを閉じ込めないまま能動的にループだけを押し出し続けます[7]。同じコヒーシンが、状況に応じて「接着」と「ループ形成」を使い分けている点が、この分子の巧妙なところです。
4. CTCFと転写マシナリー:ループの「境界」はどう決まるのか
コヒーシンがどこまでもDNAを手繰り寄せてしまっては、ゲノムの区画(TAD)はできません。ループ押し出しは、ゲノム上の特定の「障壁」にぶつかることで、きちんと境界が引かれます[8]。もっとも代表的な障壁が、DNA結合タンパク質であるCTCFです。
CTCFは、DNA上の特定の目印に「向き」を持って結合します。押し出されてきたコヒーシンが、CTCFと正面から向き合う方向(収束方向)のときにだけ、コヒーシンはそこで足を止め、安定したループ(TADの境界)が形成されます。これは単なる物理的な衝突ではなく、CTCFの特定のアミノ酸がコヒーシンの部品と立体的に結合することで達成されます。この結合部位に変化を加えると、コヒーシンはCTCFを素通りしてしまい、境界が消えてしまうことが確かめられています。
💡 用語解説:CTCFと「収束方向」
CTCFは、ゲノムの区画を仕切る「しおり」のようなタンパク質です。DNA上の目印には向きがあり、コヒーシンが押し出してくる方向とCTCFの向きが正面衝突する組み合わせ(収束方向)のときだけ、コヒーシンはそこで止まります。この仕組みのおかげで、ゲノムは規則正しい区画に整理され、遺伝子とスイッチの余計な混線が防がれます。
もう一つの障壁が、遺伝子を読み取りながらDNA上を進むRNAポリメラーゼII(転写の機械)です。この機械が進むとき、コヒーシンは前方へ押しやられ、ループが無秩序に伸びるのを防ぎます。実験的に転写の機械を細胞から取り除くと、コヒーシンへの障壁が消えて通常よりずっと長いループができ、ゲノムの接触パターンが組み替わることが示されています[8]。ループ押し出しは、CTCFと転写という2つのブレーキによって、精密にかたちづくられているのです。
5. アセチル化サイクルとWAPL:コヒーシンの「滞在時間」を決める化学スイッチ
コヒーシンが染色体にどれだけ長く留まるかは、SMC3という部品の特定の場所に「アセチル基」という小さな目印を付けたり外したりする「アセチル化サイクル」によって決まります[11]。目印を付ける酵素がESCO1とESCO2、外す酵素がHDAC8です。
💡 用語解説:アセチル化(あせちるか)
タンパク質に「アセチル基」という小さな化学の目印を付ける反応です。目印の有無で、タンパク質のはたらきやDNAとのくっつきやすさが切り替わります。コヒーシンでは、SMC3にアセチル化の目印が付くと、外す係(WAPL)から守られてDNAに長く留まれるようになり、接着が安定します。付けたり外したりのオン・オフが、コヒーシンの居場所と寿命を細かく調節しています。
ESCO1は細胞周期を通じて常にはたらき、ゲノム全体のコヒーシンやCTCFの結合部位に広く分布して、間期のループやドメインが伸びすぎないよう「制限」する調整役を担います[9]。一方のESCO2はDNA複製の時期にだけ現れ、複製の機械と強く連携して、複製したばかりの姉妹染色体を正確に接着させます[10]。このESCO2のはたらきが失われると、のちに述べるロバーツ症候群が起こります。
目印を外すHDAC8は、分裂の終わりに、切り離されて使い終わったコヒーシンからアセチル基を外し、次のサイクルで再び使えるよう「初期化」します[11]。この初期化がうまくいかないと、コヒーシンの再利用が滞ります。HDAC8の変化がコルネリア・デ・ランゲ症候群の一因になるのは、このリサイクルの障害が背景にあります。
WAPLによる「入れ替え」が細胞の個性を守る
コヒーシンは一度DNAに載っても固定されるわけではなく、WAPLによって外され、また載り直すという「入れ替え(ターンオーバー)」を絶えず繰り返しています[12]。WAPLを働かなくすると、コヒーシンが染色体上に固定されて外れなくなり、染色体が異常に凝縮した「バーミセリ(パスタ状)染色体」になります。意外なことに、コヒーシンを無理に安定化させると、かえって細胞の個性を決める遠くのスイッチ領域からコヒーシンが消え、その細胞タイプ特有の遺伝子プログラムが崩れてしまいます。つまり、WAPLが絶えずコヒーシンを解放して「自由なプール」をつくり、必要な場所へ配り直す入れ替えこそが、細胞の分化を支えているのです。この配り直しの目印役として、OCT4やSOX2といったパイオニア転写因子が関わっています。

6. 減数分裂におけるコヒーシン:2段階の分裂を成功させる巧妙な制御
卵子や精子をつくる減数分裂では、染色体の数を半分にするために、続けて2回の分裂(第一分裂・第二分裂)を行います。このためコヒーシンは、通常の分裂とはひと味違う、非常に手の込んだ制御を実装しています[13]。減数分裂では、RAD21の代わりにRec8という減数分裂専用のクレイシンが使われます。
第一分裂の準備として、相同染色体どうしを正しく対合させるため、染色体の腕の部分では、切断を伴わない「前期解離経路」によってコヒーシンがそっと外されます。哺乳類ではこの過程が、生殖細胞に多いNEK1というキナーゼを中心とした精密な調節を受けています[14]。NEK1がうまく働かないと、腕のコヒーシンが異常に残ってしまい、その後の相同染色体の分配が大きく妨げられます。
シュゴシンによる「接着の保護」と、その解除
第一分裂で相同染色体が引き離されるとき、姉妹染色体どうしはセントロメア領域では結合を保っていなければなりません。この保護を担うのがシュゴシン(Sgo1)です。シュゴシンはPP2Aという脱リン酸化酵素をセントロメアに呼び込み、セパラーゼによる切断のきっかけとなるリン酸化をすぐに打ち消すことで、セントロメアのコヒーシンを鉄壁に守ります[15]。
💡 用語解説:セパラーゼとシュゴシン
セパラーゼは、分裂の後半にクレイシン(RAD21やRec8)を切断して、姉妹染色体を切り離す「はさみ」の酵素です。シュゴシンは、その「はさみ」からセントロメアのコヒーシンを守る「ボディガード」です。減数分裂では、腕のコヒーシンは第一分裂で外し、セントロメアのコヒーシンはシュゴシンが第二分裂まで守る——というように、場所ごとに外すタイミングをずらすことで、2段階の分裂が正確に進みます。
続く第二分裂では、姉妹染色体を今度こそ両極へ引き離すため、この保護が能動的に解除されます(除保護)。第二分裂に入ると、APC/C-Cdc20という酵素が活性化してシュゴシンを分解へと追いやり、PP2Aもセントロメアから外れます[16]。守りを失ったセントロメアのRec8は一斉にリン酸化され、セパラーゼによって切断されて、姉妹染色体の最終的な二極分離が完了します。この「守って、頃合いをみて外す」という時間差の制御こそが、減数分裂の正確さを支えています。加齢に伴ってこの接着がほころびると、染色体の分配ミス(染色体異数性)が増えることも知られています。
7. コヒーシノパチー:コヒーシンの異常が起こす先天性疾患と遺伝診療
🔍 関連ページ:コルネリア・デ・ランゲ症候群1型/新生突然変異(de novo変異)/遺伝形式
生まれつき(生殖細胞系列)でコヒーシン関連遺伝子に不完全な機能低下型の変化があると、複数の臓器にまたがる先天性の症候群「コヒーシノパチー」が起こります[24]。特徴的なのは、完全にはたらきを失うのではなく(それは生命の維持に関わります)、はたらきがわずかに低下する程度の「減能型」の変化である点です。少しだけ足りない、という絶妙なバランスの崩れが、発達全体に影響します。
もっとも頻度が高いコルネリア・デ・ランゲ症候群では、NIPBLの変化が約6割を占めます。変化の種類によって重症度に傾向があり、タンパク質が途中で途切れてしまうナンセンス変異やフレームシフト変異は重症になりやすく、アミノ酸が1つ置き換わるミスセンス変異は比較的軽症になりやすい傾向が知られています。またSMC1AとHDAC8はX染色体上にあり、原因遺伝子によって遺伝形式が異なる点も、遺伝カウンセリングでは重要になります。
💡 用語解説:新生突然変異(de novo変異)
両親のどちらも持っていないのに、精子や卵子ができる過程、あるいは受精のごく初期に、お子さんで初めて新しく生じる遺伝子の変化を新生突然変異(de novo変異)といいます。コヒーシノパチーの多くはこのタイプで、家族歴がなくても起こります。このため、原因が確定している場合には、出生前の確定的な遺伝学的検査という選択肢も、ご家族が状況を理解したうえで検討できます。くわしくは新生突然変異の解説をご覧ください。
なおロバーツ症候群は、ESCO2がはたらきを失うことで、SMC3のアセチル化ができなくなり、WAPLを抑える因子を呼び込めなくなって、セントロメア付近の姉妹染色体接着が崩れる病気です[23]。またSTAG2はX染色体上にあるため、X染色体不活化のパターンによって、女性では体の部位ごとに影響の出方が異なるモザイク状の表現型を示すことがあります。診断が確定したご家族には、遺伝形式・再発の可能性・次のお子さんへの対応などを含めた、ていねいな遺伝カウンセリングが大切になります。
8. がんとコヒーシン:STAG2という「がん抑制因子」
コヒーシンは、体細胞(生まれたあとの体の細胞)でも重要ながんの舞台になります。がんゲノムの網羅的な解析から、コヒーシン関連の部品は、ヒトのがんでもっとも頻繁に変化する遺伝子群の一つであることがわかっています[17]。なかでもX染色体上にあるSTAG2は失われる変化が特に多く、膀胱がん・ユーイング肉腫・急性骨髄性白血病(AML)や骨髄異形成症候群(MDS)・膠芽腫など、幅広いがんでがん抑制因子として不活化されています。
かつては、コヒーシンの変化ががんを起こす仕組みは「接着不全による染色体の分配ミス」だと単純に考えられていました。ところが、コヒーシン変化を持つAMLや膀胱がんの多くが、染色体数の正常な細胞であることがわかり、現在では「ゲノムの3D構造が変化して、その細胞にふさわしい遺伝子プログラムが乱れる」という考え方が主流になっています[19]。
膠芽腫では、STAG2が失われると代わりにSTAG1への依存が高まり、正常な細胞には見られない極めて巨大な異常ループが多数できることが報告されています[20]。肺腺がんではSTAG2がPAXIP1/PAGR1と物理的に組み、ホルモン受容体による抗腫瘍プログラムの足場になっていることが示されました[21]。神経芽腫では、STAG2ががん遺伝子MYCNと連携しており、STAG2が減るとがん細胞が分化へと追い込まれることが分かってきています[22]。
💡 用語解説:合成致死(ごうせいちし)
2つの遺伝子のうち、片方が欠けても細胞は生きられるのに、両方そろって欠けると細胞が死んでしまう関係を「合成致死」といいます。STAG2を失ったがん細胞は、残ったSTAG1に頼って生きています。そこでSTAG1のはたらきを狙って止めると、がん細胞だけが行き詰まって死に、STAG2が正常な健康な細胞は生き残ります。副作用を抑えつつがんを狙い撃つ精密医療の考え方として、研究が進んでいます。
この「STAG2を失ったがんは、残されたSTAG1に生存を依存している」という性質を逆手に取ったのが、STAG1とSTAG2の合成致死を利用する治療戦略です[18]。正常な細胞への影響を抑えながら、がん細胞だけを狙い撃つアプローチとして開発が進められています。また、STAG2欠損のがんでDNA修復に弱点が生じる場合には、DNA修復の異常を突く薬剤への感受性も研究されています。なお、これらは研究段階の知見であり、本稿は文献に基づく学術的な解説です。
9. よくある誤解
誤解①「コヒーシンは染色体を分けるだけの糊」
接着は大切な役割ですが、それだけではありません。間期にはDNAをループ状に整理して遺伝子の読み方(発現)を制御しています。むしろ、この制御の乱れが発達障害やがんの主因と考えられるようになっています。
誤解②「コヒーシンとコンデンシンは同じもの」
よく似たリング構造を持ちますが、別の複合体です。コヒーシンは接着とループ形成、コンデンシンは分裂期の染色体凝縮と、役割が異なります。同じ設計図を土台に部品を替えて機能を分けている点が、SMCファミリーの特徴です。
誤解③「コヒーシノパチーは必ず親から遺伝する」
多くは新生突然変異(de novo変異)で、両親に同じ変化がないことが大半です。家族歴がなくても起こり得ます。原因遺伝子によって遺伝形式が違うため、再発の可能性は個別の評価が必要です。
誤解④「STAG2が壊れると染色体数がすぐ異常になる」
STAG2変化を持つがんの多くは、実は染色体数が正常です。がん化の主因は染色体数の異常ではなく、ゲノムの3D構造が変わって遺伝子プログラムが乱れることだと考えられています。
よくある質問(FAQ)
🏥 染色体・遺伝性疾患のご相談
コルネリア・デ・ランゲ症候群など
コヒーシン関連疾患を含む遺伝性疾患の検査・遺伝カウンセリングは
臨床遺伝専門医が在籍するミネルバクリニックにお気軽にご相談ください。
参考文献
- [1] Cohesin Complex: Structure and Principles of Interaction with DNA. PubMed. [PubMed 38831498]
- [2] Structural Insights into Ring Formation of Cohesin and Related Smc Complexes. PMC. [PMC4989898]
- [3] Cohesin regulation and roles in chromosome structure and function. PMC. [PMC10947815]
- [4] Cohesin in 3D: development, differentiation, and disease. Genes & Development. [Genes Dev]
- [5] Cohesin mediates DNA loop extrusion by a “swing and clamp” mechanism. PubMed. [PubMed 34624221]
- [6] High-speed AFM imaging reveals DNA capture and loop extrusion dynamics by cohesin-NIPBL. PMC. [PMC10656236]
- [7] A Brownian ratchet model for DNA loop extrusion by the cohesin complex. eLife. [eLife 67530]
- [8] Attraction and disruption: how loop extrusion and compartmentalisation shape the nuclear genome. PMC. [PMC11190842]
- [9] Cohesin recruits the Esco1 acetyltransferase genome wide to repress transcription and promote cohesion in somatic cells. PNAS. [PNAS]
- [10] Esco1 and Esco2 regulate distinct cohesin functions during cell cycle progression. PNAS. [PNAS]
- [11] Biochemical and Structural Characterization of HDAC8 Mutants Associated with Cornelia de Lange Syndrome Spectrum Disorders. Biochemistry (ACS). [ACS]
- [12] WAPL maintains a cohesin loading cycle to preserve cell-type specific distal gene regulation. PMC. [PMC7610352]
- [13] Meiosis-specific prophase-like pathway controls cleavage-independent release of cohesin by Wapl phosphorylation. PLOS Genetics. [PLOS Genetics]
- [14] Cohesin removal along the chromosome arms during the first meiotic division depends on a NEK1-PP1γ-WAPL axis in the mouse. PMC. [PMC5123770]
- [15] Shugoshin collaborates with protein phosphatase 2A to protect cohesin. PubMed. [PubMed 16541025]
- [16] APC/C-Cdc20 mediates deprotection of centromeric cohesin at meiosis II in yeast. PMC. [PMC5499901]
- [17] Cohesin Mutations in Human Cancer. PMC. [PMC4980180]
- [18] Synthetic lethality between the cohesin subunits STAG1 and STAG2 in diverse cancer contexts. PMC. [PMC5531830]
- [19] Cohesin mutations in myeloid malignancies made simple. PMC. [PMC6601335]
- [20] STAG2 mutations regulate 3D genome organization, chromatin loops, and Polycomb signaling in glioblastoma multiforme. PubMed. [PubMed 38705393]
- [21] PAXIP1 and STAG2 converge to maintain 3D genome architecture and facilitate promoter/enhancer contacts to enable stress hormone-dependent transcription. PubMed. [PubMed 37070193]
- [22] Stag2-cohesin mediates 3D genome dynamics to regulate MYCN driven oncogenic transcriptome in neuroblastoma. Cancer Research (AACR). [AACR]
- [23] The molecular mechanism underlying Roberts syndrome involves loss of ESCO2 acetyltransferase activity. Human Molecular Genetics. [Oxford HMG]
- [24] Cornelia de Lange syndrome: from molecular diagnosis to therapeutic approach. PMC. [PMC7231464]






