KHドメインは、ヒトの異質核リボ核酸タンパク質(hnRNP)Kで初めて見つかったタンパク質ドメインで、進化的に保存された約70アミノ酸から成る配列です。このドメインは、さまざまな核酸結合タンパク質に存在し、RNAと結合してRNAを認識する機能を持っています。一部のタンパク質では、複数のKHドメインが存在し、それらが協力することも、単独で機能することもあります。KHドメインタンパク質であるNOVA1に対する自己抗体は、腫瘍随伴性オプソクローヌス(リズム、方向、振幅すべてが不規則な衝動的な眼球運動が絶え間なく発生する状態)を引き起こすことがあります。また、KHドメインはリボソームタンパク質S3のN末端にも存在します。
KHドメインの構造
KHドメインは、RNA結合タンパク質に見られる構造モチーフであり、約70アミノ酸から構成されています。このドメインは、RNAとの結合に関与し、RNAの安定性、翻訳、転写後修飾などの生物学的プロセスにおいて重要な役割を果たします[5][7]。
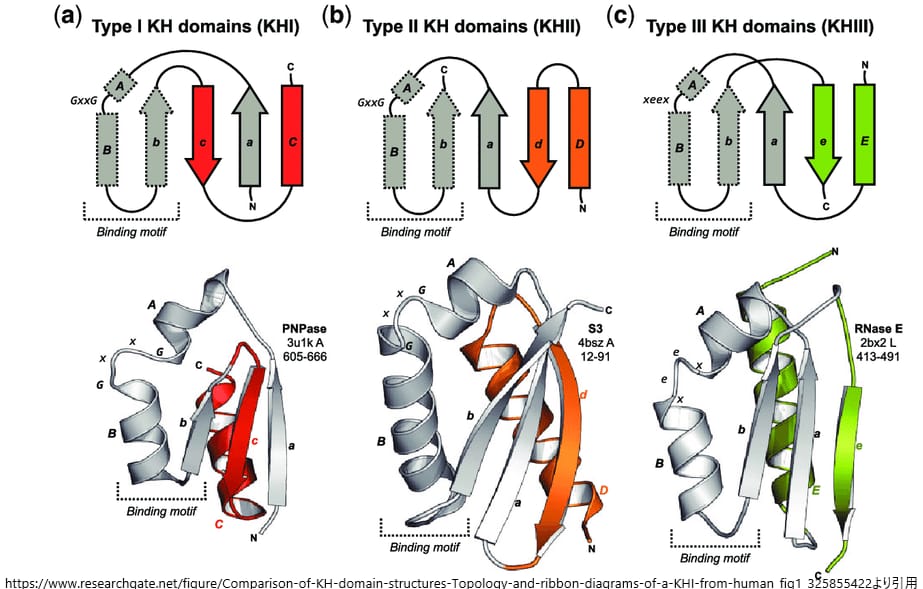
KHドメインには、タイプIとタイプIIの2種類の構造が存在することが明らかにされています。タイプIは主に真核生物のタンパク質に存在し、タイプIIは主に原核生物のタンパク質に見られます[5]。これらのドメインは、RNAとの特異的な結合を可能にするための特定の構造的特徴を持っています。
タイプI KHドメインは、βシートとαヘリックスからなる特徴的な折りたたみを持ち、この構造がRNAとの結合面を形成します。一方、タイプII KHドメインは、タイプIとは異なる折りたたみを持ち、RNAとの結合において異なる機構を持つ可能性があります[5]。
具体的なKHドメインの構造については、研究によってさまざまなKHドメインを含むタンパク質の結晶構造が解析されています。例えば、リボソーム成熟に関わるRbfAタンパク質の構造解析により、RbfAがKHドメイン構造を有していることが判明しました[1]。このような構造解析は、KHドメインがRNAとどのように相互作用するかを理解する上で重要な情報を提供しています。
また、KHドメインは、RNA結合において重要な役割を果たす他のドメインと共に存在することがあります。例えば、FMRP(Fragile X Mental Retardation Protein)は、3つのKHドメインと1つのRGGボックスを含むことが知られており、これらのドメインがRNAとの結合に関与しています[2]。
これらの知見は、KHドメインがRNAとの結合においてどのように機能するかを理解するための基盤を提供し、RNA結合タンパク質の機能解明や疾患メカニズムの理解に貢献しています。
- 参考文献・出典
-
[1] https://www.riken.jp/medialibrary/riken/pr/press/2007/20071109_1/20071109_1.pdf
[2] https://www.jstage.jst.go.jp/article/fpj/154/6/154_294/_pdf
[3] https://bsd.neuroinf.jp/wiki/RNA%E7%B5%90%E5%90%88%E3%82%BF%E3%83%B3%E3%83%91%E3%82%AF%E8%B3%AA
[4] https://www.aist.go.jp/aist_j/press_release/pr2012/pr20120910/pr20120910.html
[5] https://ja.wikipedia.org/wiki/KH%E3%83%89%E3%83%A1%E3%82%A4%E3%83%B3
[6] https://ja.wikipedia.org/wiki/.kh
[7] https://www.jstage.jst.go.jp/article/membrane/34/3/34_133/_pdf
KHドメインの機能
KHドメインは、RNAや一本鎖DNAに結合する能力を持っています。このドメインにおいて、核酸はドメインの片側で伸びた状態で結合します。この結合は、αヘリックス1、αヘリックス2、高度に保存された配列モチーフを含むGXXGループ、そして可変ループの間に形成されるクレフトで起こります。結合クレフトは疎水性であり、複合体を安定化させるために様々なタンパク質固有の相互作用が追加されています。この構造的配置により、KHドメインは核酸との効果的な結合を実現し、その生物学的機能を果たしています。
(参考文献)
KHドメイン(K homology domain)は、約70アミノ酸からなるRNA結合ドメインの一種で、RNAに結合して認識する機能を持ちます。このドメインは、ヒトhnRNP Kタンパク質で最初に同定されたことからKHと命名されましたが、バクテリアから哺乳類に至るまで広く存在しています。KHドメインを含むタンパク質は、RNAと結合することで、RNAの安定性、翻訳、転写後修飾など、RNAの様々な生物学的プロセスに関与しています[1][2][5]。
代表的なKHドメインを含むタンパク質には、脆弱X症候群の原因遺伝子産物であるFMRPや、AU-rich element(ARE)結合タンパク質KSRPなどがあります。これらのタンパク質は、RNAとの結合を通じて、細胞内でのRNAの機能を調節する重要な役割を担っています[1]。
さらに、いくつかのタンパク質には複数のKHドメインが存在し、それらが協働的に機能することで、RNAとの結合の特異性や親和性を高め、RNAに対する調節機能を強化しています[2][5]。KHドメインは、RNA結合タンパク質の中でも特に広範囲の核酸結合タンパク質に存在しており、細胞内でのRNA関連プロセスにおいて中心的な役割を果たしていることが示されています[2][5]。
- 参考文献・出典
-
[1] https://www.yodosha.co.jp/jikkenigaku/keyword/1992.html
[2] https://www.weblio.jp/content/KH%E3%83%89%E3%83%A1%E3%82%A4%E3%83%B3
[3] https://www.bi.a.u-tokyo.ac.jp/~shimizu/seq3.pdf
[4] https://bsd.neuroinf.jp/wiki/RNA%E7%B5%90%E5%90%88%E3%82%BF%E3%83%B3%E3%83%91%E3%82%AF%E8%B3%AA
[5] https://ja.wikipedia.org/wiki/KH%E3%83%89%E3%83%A1%E3%82%A4%E3%83%B3
KHドメインの種類
タイプIドメインは主に真核生物のタンパク質に見られ、タイプIIドメインは主に原核生物のタンパク質に存在します。これらのドメインは最小限のコンセンサス配列モチーフを共有していますが、構造の折りたたみ方に違いがあります。タイプIのKHドメインは3本鎖のβシートから構成されており、これらはすべて反平行です。一方、タイプIIドメインでは、3本のβシートのうち2本が平行に配置されています。また、タイプIドメインは一般的に同じタンパク質内に複数存在することが多いのに対し、タイプIIドメインは通常、一つのタンパク質に1個だけ存在します。







