目次
EF-hand domainとは
EF-handドメインは、タンパク質の二次構造のモチーフの一つであり、約40アミノ酸残基から成り立っています。このモチーフは、ヘリックス-ループ-ヘリックスの構造を持ち、細胞内カルシウムの結合に関与しています。EF-handドメインはしばしば一つまたは複数の対をなして存在し、それがEF-handモチーフを含むタンパク質に様々な構造上・機能上の多様性をもたらしています。EF-handを含むタンパク質は、機能的に調節的グループと構造的グループに分類することができます。カルシウムが調節的なEF-handドメインを含むタンパク質に結合すると、立体構造の変化をもたらし、それがしばしば酵素反応を伴いながら標的タンパク質に伝達されます。これと対照的に、カルシウムが構造的なEF-handドメインを含むタンパク質に結合しても、特に立体構造の変化は起こしません。構造的なEF-handドメインは、細胞内カルシウム量を緩衝する役割を持つ可能性があります[2][3]。
- 参照・引用
-
[2] https://www.cellsignal.jp/learn-and-support/protein-domains-and-interactions/efhand-protein-domain
[3] https://ja.wikipedia.org/wiki/EF%E3%83%8F%E3%83%B3%E3%83%89
EF-handドメインを含むタンパク質の例には以下のものがあります:
– カルモジュリン(細胞内のカルシウムシグナル伝達に関与)
– トロポニンC(筋収縮の調節)
– S100タンパク質(細胞内プロセスの調節)
– パルモジュリン関連タンパク質
EF-hand domainの名前の由来
EFハンドドメインの名前は、その形状が英文字の「E」と「F」に似ていることに由来しています。具体的には、このドメインは、2つのαヘリックス(らせん状の構造)が短いループでつながれており、このループがカルシウムイオン(Ca2+)を結合する部分となっています。この構造が、1960年代後半にX線結晶構造解析によって初めて観察されたとき、構造生物学者たちは、この特徴的な形状が、特定のタンパク質の3次元構造の図表で使われる英文字に似ていることに注目しました。そこで、このモチーフが最初に詳細に記述されたパルバルブミンの構造内で、この特徴的なドメインが位置していた二つのヘリックスが「E」ヘリックスと「F」ヘリックスと名付けられ、その間にあるループに由来して「EFハンド」という名前がつけられました。この名称は、その後、カルシウムを結合する同様の構造を持つ他のタンパク質のドメインにも適用されるようになりました。
EF-hand domainの構造
EFハンドドメインは、タンパク質の二次構造のモチーフの一つで、主にカルシウムイオンを結合する機能を持っています。このモチーフは、互いにおよそ垂直になっている2つのαヘリックスから構成されており、その間にループ状の構造が存在します。このループがカルシウムイオンと結合する部分であり、12個のアミノ酸残基から成るカルシウム結合ループが特徴的です[4]。
EFハンドモチーフは約40アミノ酸残基から成り、細胞内カルシウムの結合に関与しています。EFハンドドメインはしばしば1つまたは複数の対をなして存在し、それがEFハンドモチーフを含むタンパク質に様々な構造上・機能上の多様性をもたらしています。EFハンドを含むタンパク質は機能的に2つの区分、すなわち調節的グループと構造的グループに分類することができます。カルシウムが調節的なEFハンドドメインを含むタンパク質に結合すると立体構造の変化をもたらし、それがしばしば酵素反応を伴いながら標的タンパク質に伝達されます。これと対照的に、カルシウムが構造的なEFハンドドメインを含むタンパク質に結合しても、特に立体構造の変化は起こしません。構造的なEFハンドドメインは、おそらく細胞内カルシウム量を緩衝する役割を持ちます[3]。
また、EFハンドカルシウム結合タンパク質は、ヘリックスループヘリックス構造であるEFハンドが動物ゲノムに見られる最も一般的なモチーフの1つであり、EFハンドCa(2+)結合タンパク質(EFCaBPs)は細胞全体に広く分布しています。しかし、研究者は、ペプチド配列が特定の機能をコードする方法や、これらの知識が健康と病気の中でEFCaBPsの役割を定義し、最終的にはCa(2+)の制御や設計さえ可能にする方法を理解することによって困惑しています[1]。
EFハンド様のタンパク質は、細菌においても発見されており、これらのプロカリオートEFハンド様タンパク質はカルシウムシグナリングと恒常性に広く関与しています。これらはEFハンドモチーフと異なる柔軟な長さのカルシウム結合ループを含んでいますが、その調整特性は古典的なEFハンドモチーフに似ています[2]。
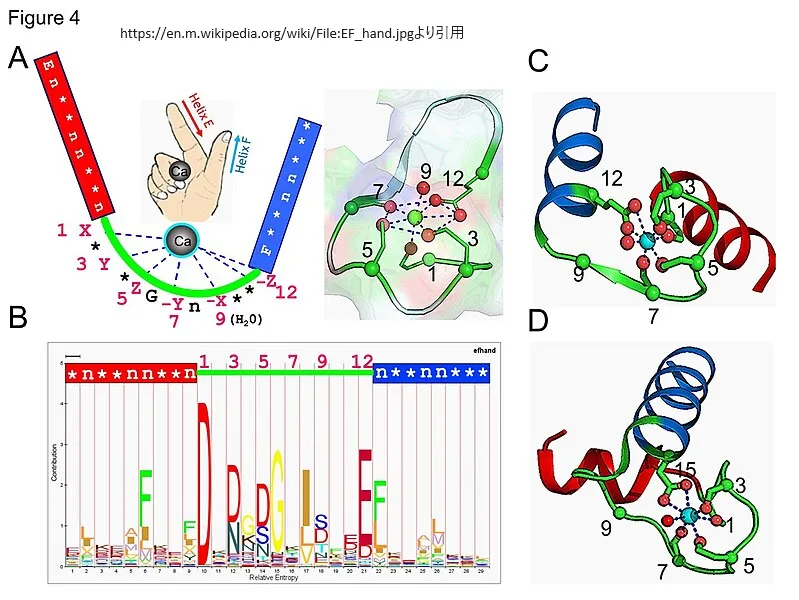
https://en.m.wikipedia.org/wiki/File:EF_hand.jpg
より引用
(A)、標準的なEF-ハンドCa2+結合モチーフの模式図。EF-handモチーフは、ヒトの手の親指と人差し指を広げたような29残基のらせん-ループ-らせんトポロジーを含んでいます。Ca2+は、側鎖のカルボキシル基またはヒドロキシル基(ループ配列1、3、5、12位)の7個の酸素原子、主鎖のカルボニル基(7位)、および架橋水(9位を介して)を含む12残基ループ内のリガンドによって配位されます。12位の残基は二座配位子として機能。nは疎水性残基。(B)、カノニカルEFハンドモチーフのHMMロゴ(http://pfam.sanger.ac.uk/family?acc=PF00036)。いくつかの位置でアミノ酸が保存されていることから、ゲノム配列からEFハンドモチーフを予測することが可能。(C)、カルモジュリンの典型的なカノニカルEF-ハンドモチーフの立体構造(PDBコード:3cln)。Ca2+は12残基ループのリガンドによってキレートされます。(D)、大腸菌の溶菌トランスグリコシラーゼBの可溶性断片から得られたEF-ハンド様モチーフの立体構造(PDBコード:1qut)。このモチーフは、2つの耳輪に挟まれた15残基(12残基ではなく)のCa2+結合ループを含んでいます。
- 参照・引用
-
[1] https://bibgraph.hpcr.jp/abst/pubmed/21314091?click_by=rel_abst
[2] https://en.wikipedia.org/wiki/EF_hand
[3] https://www.cellsignal.jp/learn-and-support/protein-domains-and-interactions/efhand-protein-domain
[4] https://ja.wikipedia.org/wiki/EF%E3%83%8F%E3%83%B3%E3%83%89
[5] https://www.jstage.jst.go.jp/article/jcac/7/0/7_0_78/_pdf
[6] https://jglobal.jst.go.jp/detail?JGLOBAL_ID=201202204414299921
[7] https://kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-08272201/
[8] https://bibgraph.hpcr.jp/abst/pubmed/19129176?click_by=p_ref
EF-hand domainの機能
EF-handドメインは、約40アミノ酸残基から成り、細胞内カルシウムの結合に関与しています[5]。このモチーフは、動物ゲノムに見られる最も一般的なモチーフの1つであり、EF-handカルシウム結合タンパク質(EFCaBPs)は細胞全体に広く分布しています[2]。EF-handドメインはしばしば1つまたは複数の対をなして存在し、それがEF-handモチーフを含むタンパク質に様々な構造上・機能上の多様性をもたらしています[5]。
EF-handを含むタンパク質は機能的に2つの区分、すなわち調節的グループと構造的グループに分類することができます。カルシウムが調節的なEF-handドメインを含むタンパク質に結合すると立体構造の変化をもたらし、それがしばしば酵素反応を伴いながら標的タンパク質に伝達されます。これと対照的に、カルシウムが構造的なEF-handドメインを含むタンパク質に結合しても、特に立体構造の変化は起こしません。構造的なEF-handドメインは、おそらく細胞内カルシウム量を緩衝する役割を持ちます[5]。
EF-handドメインは、カルシウム依存的な相互作用を介して、細胞内のシグナル伝達やホメオスタシスに重要な役割を果たします。例えば、カルシウム結合によってタンパク質の構造が変化し、その結果として他のタンパク質との相互作用が変わることがあります。このような機能は、細胞の様々な生理的プロセスにおいて重要です[4][5]。
また、EF-handドメインは、カルシウム結合によってタンパク質の活性化や機能の変化を引き起こすことが知られています。例えば、カルモジュリンやS-100タンパク質などが、カルシウム結合によって様々なタンパク質と相互作用し、細胞内のシグナル伝達に関与しています[5]。
さらに、EF-handドメインは、カルシウム結合によって細胞内のカルシウム濃度の変化を感知し、それに応じて細胞の応答を調節する役割を果たすことが示唆されています。これは、カルシウムが細胞内の第二メッセンジャーとして機能することと関連しています[2][4][5]。
総じて、EF-handドメインは、カルシウム結合による構造変化を介して、細胞内の多様な生理的プロセスを調節する重要な役割を担っています。
- 参照・引用
-
[1] https://pdbj.org/emnavi/quick.php?id=pdb-2z14&lang=ja
[2] https://bibgraph.hpcr.jp/abst/pubmed/21314091?click_by=rel_abst
[3] https://seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2019.910191/index.html
[4] https://en.wikipedia.org/wiki/EF_hand
[5] https://www.cellsignal.jp/learn-and-support/protein-domains-and-interactions/efhand-protein-domain
[6] https://www.jstage.jst.go.jp/article/jcac/7/0/7_0_78/_pdf
[7] https://ja.wikipedia.org/wiki/EF%E3%83%8F%E3%83%B3%E3%83%89
[8] https://bibgraph.hpcr.jp/abst/pubmed/7685034?click_by=p_ref
EF-hand domainの機能不全
EF-hand domainは、カルシウムイオンを結合することで知られるタンパク質の二次構造モチーフです。このドメインは、細胞内カルシウムの結合に関与し、カルシウムシグナリングやホメオスタシスに重要な役割を果たします。EF-handドメインを含むタンパク質は、調節的グループと構造的グループに分類され、カルシウムが調節的なEF-handドメインを含むタンパク質に結合すると立体構造の変化をもたらし、標的タンパク質に伝達されることがしばしばあります。これに対して、カルシウムが構造的なEF-handドメインを含むタンパク質に結合しても、特に立体構造の変化は起こしませんが、細胞内カルシウム量を緩衝する役割を持つとされています[6]。
EF-hand domainの機能不全は、カルシウム結合能力の低下やタンパク質の立体構造の変化により、カルシウムシグナリングの誤調節を引き起こす可能性があります。これは、細胞の機能不全や疾患の発症につながる可能性があります。例えば、EF-handドメインを持つカルパインは、カルシウムにより活性化され、細胞内の基質となるタンパク質を限定的に分解することで細胞機能に関わっていますが、カルパインの活性の異常な亢進や低下は、神経変性や筋ジストロフィー症などの疾患の原因となることが報告されています[10]。
また、EF-handドメインの機能不全は、カルシウム依存的な相互作用因子の異常や細胞死、刺激応答反応における役割の障害にも関連しています。特に、penta-EF-hand(PEF)ドメインを持つタンパク質ファミリーは、細胞死や刺激応答反応における重要な役割を果たしており、このドメインの機能不全は、これらのプロセスの誤調節につながる可能性があります[11]。
EF-handドメインの機能不全による影響は、カルシウム結合能力の低下やタンパク質の立体構造の変化によるものであり、これによりカルシウムシグナリングや細胞機能の誤調節が引き起こされる可能性があります。これは、細胞の機能不全や疾患の発症につながる可能性があるため、EF-handドメインの正常な機能は細胞の健康と疾患の予防において重要です。
- 参照・引用
-
[6] https://www.cellsignal.jp/learn-and-support/protein-domains-and-interactions/efhand-protein-domain
[10] https://www.tmig.or.jp/J_TMIG/genome300/CAPN1.html
[11] https://www.jstage.jst.go.jp/article/kagakutoseibutsu1962/38/12/38_12_792/_pdf
EF-hand domainをターゲットとした研究開発
EF-handドメインは、カルシウムイオンを結合する能力を持つタンパク質構造の一種であり、多様な生物学的プロセスにおいて重要な役割を果たしています。このドメインは、カルシウム結合タンパク質に特有の構造ドメインまたはモチーフであり、ヘリックス-ループ-ヘリックス構造を持ちます。EF-handドメインを含むタンパク質は、カルシウムイオンの結合に応じてその機能や構造を変化させることができ、細胞内カルシウムシグナリングやカルシウム依存的な様々な生物学的プロセスに重要な役割を果たします[1]。
EF-handドメインをターゲットとした研究開発は、このドメインの構造と機能の理解を深めることを目的としています。例えば、EF-handドメインを含むタンパク質の構造解析により、カルシウム結合時の立体構造の変化や、カルシウム結合によるタンパク質の活性化メカニズムの解明が進められています[2][4]。また、EF-handドメインを含むタンパク質の機能解析により、細胞内カルシウムシグナリングにおける役割や、カルシウム依存的な生物学的プロセスへの関与が研究されています[3][5]。
さらに、EF-handドメインを含むタンパク質は、疾患の発症メカニズムに関与していることが示唆されており、これらのタンパク質をターゲットとした治療薬の開発も進められています。例えば、グアニンヌクレオチド交換因子SLATのEF-handドメインおよびPHドメインとIP3受容体1との会合がT細胞におけるCa2+シグナル伝達を促進することが示され、この相互作用が選択的免疫抑制薬の標的となりうることが示唆されています[9]。
このように、EF-handドメインをターゲットとした研究開発は、基礎研究から応用研究に至るまで幅広く行われており、生命科学のみならず医学や薬学の分野においても重要な意義を持っています。
- 参照・引用
-
[1] https://ejje.weblio.jp/content/ef%E3%83%8F%E3%83%B3%E3%83%89%E3%83%89%E3%83%A1%E3%82%A4%E3%83%B3
[2] https://www.cellsignal.com/learn-and-support/protein-domains-and-interactions/efhand-protein-domain
[3] https://www.guidetopharmacology.org/GRAC/FamilyDisplayForward?familyId=915
[4] https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2373453/
[5] https://en.wikipedia.org/wiki/EF_hand
[6] https://www.cellsignal.jp/learn-and-support/protein-domains-and-interactions/efhand-protein-domain
[8] https://bsd.neuroinf.jp/wiki/%E3%82%AB%E3%83%AB%E3%83%A2%E3%82%B8%E3%83%A5%E3%83%AA%E3%83%B3
[9] https://www.cosmobio.co.jp/aaas_signal/archive/ra-20140930-3.asp
EF-hand domain containingに属する遺伝子
AIF1
AIF1L
ANKEF1
CABP1
CABP2
CABP4
CABP5
CABP7
CACNA1B
CACNA1E
CALB1
CALB2
CALML3
CALML4
CALML5
CALML6
CALM1
CALM2
CALM3
CALN1
CAPNS1
CAPNS2
CAPN1
CAPN2
CAPN3
CAPN8
CAPN9
CAPN11
CAPN12
CAPN14
CAPS
CAPSL
CAPS2
CETN1
CETN2
CETN3
CGREF1
CHP1
CHP2
CIB1
CIB2
CIB3
CIB4
CRACR2A
CRACR2B
CRNN
DGKA
DGKB
DGKG
DNAH7
DST
DUOX1
DUOX2
CLXN
EFCAB2
EFCAB3
EFCAB5
EFCAB6
EFCAB7
EFCAB8
EFCAB9
EFCAB10
EFCAB11
EFCAB12
EFCAB13
EFCAB14
EFCC1
EFHB
EFHC1
EFHC2
EFHD1
EFHD2
EHD1
EHD2
EHD3
EHD4
EPS15
EPS15L1
FKBP7
FKBP9
FKBP10
FKBP14
FLG
FLG2
FSTL4
FSTL5
GCA
GNPTAB
GPD2
GUCA1A
GUCA1B
GUCA1C
HPCA
HPCAL1
HPCAL4
HRNR
ITSN1
ITSN2
KCNIP1
KCNIP2
KCNIP3
KCNIP4
LCP1
LETM1
LETM2
LPCAT1
LPCAT2
MACF1
MCC
MCFD2
MICU1
MICU2
MICU3
NCALD
NCS1
NECAB1
NECAB2
NECAB3
NIN
NINL
NKD1
NKD2
NOX5
NUCB1
NUCB2
OCM
OCM2
PDCD6
PEF1
PKD2
PLCD1
PLCD4
PLCG1
PLCH1
PLCH2
PLCZ1
PLS1
PLS3
PPEF1
PPEF2
PPP2R3A
PPP2R3B
PPP3R1
PPP3R2
PRKCSH
PVALB
RAB11FIP3
RAB11FIP4
RASEF
RASGRP1
RASGRP2
RASGRP3
RASGRP4
RCVRN
REPS1
REPS2
RHBDL3
RHOT1
RHOT2
RPTN
RYR2
RYR3
SCGN
SELENON
SLC25A12
SLC25A13
SLC25A23
SLC25A24
SLC25A25
SPATA21
SPTAN1
SPTA1
SRI
SWAP70
S100A1
S100A2
S100A4
S100A5
S100A6
S100A7
S100A7A
S100A7L2
S100A8
S100A9
S100A11
S100A12
S100A16
S100B
S100G
S100P
S100Z
TBC1D8B
TBC1D9
TBC1D9B
TCHH
TESC
TNNC1
TNNC2
USP32
VSNL1
ZZEF1







