目次
この記事では、Dblファミリーのグアニンヌクレオチド交換因子の機能と、Rho GTPasesの活性化における役割について解説します。生化学的プロセスにおけるこれらの因子の重要性と、疾患感受性に及ぼす影響についても探求します。
第1章: Dblファミリーの概要
Dblファミリーの定義と特徴
Dblファミリーは、グアニンヌクレオチド交換因子(GEF)の一種で、GTP結合タンパク質(GTPアーゼ)の活性化を促進するタンパク質またはタンパク質ドメインです。これらの交換因子は、GTPアーゼ上に結合しているグアノシン二リン酸(GDP)の放出を促し、グアノシン三リン酸(GTP)の結合を促進することで、GTPアーゼの活性化を行います。Dblファミリーは、その構造と機能において、他のグアニンヌクレオチド交換因子とは異なる特徴を持っています。
Dblファミリーの特徴としては、DH(Dbl homology)ドメインを持つことが挙げられます。このDHドメインは、GTPアーゼの活性化に直接関与しており、多くの場合、隣接するPH(pleckstrin homology)ドメインと共に存在します。PHドメインは細胞膜への結合や細胞内の特定のリン脂質との相互作用に関与しています。
また、Dblファミリーのタンパク質は、組織特異的な発現を示すことが多く、哺乳類においては、細胞シグナリングの複雑さと比例してDblファミリータンパク質の数が増加していることが観察されています[6]。これは、Dblファミリーが細胞の様々なシグナリング経路において重要な役割を果たしていることを示唆しています。
DOCKファミリーはDblファミリーとは別のグループであり、DHドメインとは構造的にも配列的にも関連性がないことも特筆されます。これまでに11のDOCKファミリーのメンバーが同定されており、これらはRacなどのGTPアーゼを活性化することが知られています[6]。
グアニンヌクレオチド交換因子の役割
グアニンヌクレオチド交換因子(GEF)は、細胞内のシグナル伝達経路において重要な役割を果たすタンパク質やタンパク質ドメインです。これらはGTPアーゼと呼ばれる一群の酵素の活性化に関与しています。GTPアーゼは、細胞内で分子スイッチのような役割を担い、GDP(グアノシン二リン酸)結合型とGTP(グアノシン三リン酸)結合型の間で切り替わることによって、細胞の様々な機能を制御しています[4]。
GEFは、GTPアーゼがGDPを放出し、GTPを結合することを促進することで、GTPアーゼを活性化します。この活性化により、GTPアーゼは細胞増殖、細胞骨格の形成、細胞移動などの細胞機能を調節することができます。例えば、RhoAという低分子量Gタンパク質は、細胞骨格の形成に関与しており、GEFであるSmgGDS-558によって活性化されることが知られています[2]。
また、GEFは特定の細胞内の場所でのみ活性化することがあり、その結果、細胞内でのシグナルの局所化を可能にします。これにより、細胞はより精密に機能を調節することができます。例えば、STEFというGEFは、神経細胞の移動や神経突起の伸長、シナプスの形成など、発生過程の脳における特定の過程で強く発現していることが示されています[1]。
さらに、GEFはがんの発生や進行にも関与していることが示唆されており、GEFを標的とした抗がん剤の開発が期待されています。これは、GEFが細胞増殖や細胞移動などの過程を制御するため、これらの機能ががん細胞の成長や転移に関与しているからです[2]。
総じて、グアニンヌクレオチド交換因子は、細胞の様々な生理的プロセスを調節するために不可欠なタンパク質であり、その機能の理解は細胞生物学や医学研究において重要な意味を持っています。
- 参考文献・出典
-
[1] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-13780600/
[2] www.f.u-tokyo.ac.jp/manages/topics/data/1536299827_1.pdf
[3] www.funakoshi.co.jp/contents/68824
[4] ja.wikipedia.org/wiki/%E3%82%B0%E3%82%A2%E3%83%8B%E3%83%B3%E3%83%8C%E3%82%AF%E3%83%AC%E3%82%AA%E3%83%81%E3%83%89%E4%BA%A4%E6%8F%9B%E5%9B%A0%E5%AD%90
[5] bibgraph.hpcr.jp/abst/pubmed/24308970?click_by=rel_abst
[6] jglobal.jst.go.jp/public/200902275307670608
[7] pfwww.kek.jp/pf-sympo/24/abstract/UG03_Sato.pdf
第2章: Rho GTPasesとの関係
Rho GTPasesの基本機能
Rho GTPasesは細胞の形態や運動性を制御するために重要な役割を果たす低分子量Gタンパク質の一群です。これらのタンパク質は細胞骨格のダイナミクス、特にアクチンフィラメントの組織化に関与しており、細胞の形状変化、接着、分裂、遊走などのプロセスを調節します。
RhoファミリーGTPaseは、細胞突起の形成、伸長、退縮、分解に必要なアクチンフィラメントの動的な再編成を担っています。これには糸状仮足と葉状仮足の形成が含まれ、それぞれRacとCdc42によって調節されます[1]。細胞-ECM(細胞外マトリックス)接着や細胞遊走におけるRhoファミリーGTPasesの役割はよく研究されていますが、まだ解明されていない問題も多く残されています。2D培養から得られたデータは、現在では3D細胞培養モデルでの確認が必要とされています[1]。
Rho GTPasesの活性は、GTPの結合によって制御されます。活性化された状態では、細胞のアクチン細胞骨格の再編成を促進し、細胞の方向性応答に必要な遊走を引き起こします。この遊走は、細胞外の誘導合図に対する細胞の方向性応答であり、例えばケモカインや増殖因子などの環境合図に向かう、またはそれから離れる動きを指します[1]。
また、Rho GTPasesは細胞内のシグナル伝達経路においても中心的な役割を果たしており、細胞の生存、増殖、分化などの基本的な生物学的プロセスに影響を与えます。これらのタンパク質は、細胞内での情報伝達のハブとして機能し、多くの下流の効果器を介して細胞の応答を調節します。
Rho GTPasesの機能は、GTPのローディングだけでなく、ゲラニルゲラニル化といった翻訳後修飾にも依存しています。この修飾は細胞の局在化や機能に影響を与えることが知られています[3][4]。
低分子量Gタンパク質は、グアニンヌクレオチド交換因子(GEFs)によって活性化され、GTPase活性化タンパク質(GAPs)によって不活性化されます。これにより、細胞内でのRho GTPasesの活性状態が精密に制御されます[5]。
総じて、Rho GTPasesは細胞の形態と機能を調節するために不可欠な分子であり、細胞の動的な振る舞いを制御するための中心的な役割を担っています。
- 参考文献・出典
-
[1] www.cosmobio.co.jp/product/detail/cytoskeleton_news_202001.asp?entry_id=36603
[2] bibgraph.hpcr.jp/abst/pubmed/17680807?click_by=p_ref
[3] bibgraph.hpcr.jp/abst/pubmed/18364514?click_by=rel_abst
[4] www.cosmobio.co.jp/aaas_signal/archive/pr_20080325_3.asp
[5] ja.wikipedia.org/wiki/%E3%82%B0%E3%82%A2%E3%83%8B%E3%83%B3%E3%83%8C%E3%82%AF%E3%83%AC%E3%82%AA%E3%83%81%E3%83%89%E4%BA%A4%E6%8F%9B%E5%9B%A0%E5%AD%90
[6] www.cellsignal.jp/products/primary-antibodies/rho-gtpase-antibody-sampler-kit/9968
[7] jglobal.jst.go.jp/detail?JGLOBAL_ID=201602281848689041
DblファミリーとRho GTPasesの相互作用
Dblファミリーは、Rho GTPasesの活性を調節する重要な役割を果たしています。Rho GTPasesは細胞の形態、運動性、増殖、および生存に関与する低分子量GTP結合タンパク質であり、細胞内シグナル伝達の重要な調節因子です。Dblファミリーは、Rho GTPasesに対するグアニンヌクレオチド交換因子(GEF)として機能し、GDPからGTPへの交換を促進することで、Rho GTPasesを活性化状態に導きます。
Dblファミリーメンバーは、Dbl相同性(DH)ドメインとプレクストリン相同性(PH)ドメインを含む構造を持ち、これらのドメインがRho GTPasesの特異的な活性化に関与しています。例えば、KIAA0380はDblファミリーメンバーであり、RhoAに対して特異的なGEF活性を持つが、Rac1やCdc42には特異的ではないことが報告されています[4]。このように、DblファミリーはRho GTPasesの中でも特定のメンバーを選択的に活性化する能力を持っています。
また、Dblファミリーの活性化は、細胞内の様々なシグナルに応答して行われます。例えば、チロシンリン酸化はRhoファミリーGTPaseの活性化や結合パートナーの調節に重要な役割を果たし[1]、三量体Gタンパク質Gβγサブユニットによる制御もRho特異的グアニンヌクレオチド交換因子の活性化に関与しています[3]。
さらに、Dblファミリーは細胞のアクチン骨格の再構築にも重要な役割を果たしています。Rho GTPasesはアクチン骨格のダイナミクスを調節し、細胞の形態変化や運動性に影響を与えます。DblファミリーによるRho GTPasesの活性化は、アクチンストレスファイバーの形成や細胞接着斑の誘導など、細胞骨格の再構築を促進します[2][4]。
このように、DblファミリーはRho GTPasesの活性化を介して細胞内シグナル伝達と細胞骨格の再構築を調節し、細胞の様々な機能に影響を与える重要な役割を担っています。
- 参考文献・出典
-
[1] www.cosmobio.co.jp/product/detail/cytoskeleton_news_201605.asp?entry_id=16904
[2] bsd.neuroinf.jp/wiki/Rho%E3%83%95%E3%82%A1%E3%83%9F%E3%83%AA%E3%83%BC%E4%BD%8E%E5%88%86%E5%AD%90%E9%87%8FG%E3%82%BF%E3%83%B3%E3%83%91%E3%82%AF%E8%B3%AA
[3] seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2016.880416/data/index.html
[4] bibgraph.hpcr.jp/abst/pubmed/10526156?click_by=p_ref
第3章: 生化学的プロセスにおけるDblファミリー
細胞内シグナル伝達におけるDblファミリーの役割
細胞内シグナル伝達におけるDblファミリーの役割について解説します。Dblファミリーは、細胞内シグナル伝達において重要な役割を果たす一群のタンパク質であり、特に低分子量GTP結合タンパク質の活性化に関与しています。これらのタンパク質は、細胞の増殖、分化、運動性、形態形成など多岐にわたる生命現象を調節することが知られています。
● Dblファミリーの基本機能
Dblファミリーのタンパク質は、GTPase活性化因子(GEF)として機能し、RhoファミリーGTPaseの活性化を促進します。RhoファミリーGTPaseには、Rho、Rac、Cdc42などが含まれ、これらは細胞骨格の再構築、細胞接着、細胞運動などを制御します。DblファミリーGEFは、これらのGTPaseがGDP(不活性型)からGTP(活性型)へと結合する核酸を交換することを促進し、結果としてGTPaseの活性化を引き起こします[3][4]。
● 細胞機能への影響
DblファミリーによるRhoファミリーGTPaseの活性化は、細胞の形態変化、運動性、および細胞間相互作用に直接影響を与えます。例えば、RhoAの活性化は細胞の収縮と接着を促進し、Racの活性化は細胞の伸展と運動を促進し、Cdc42の活性化は細胞の極性形成に関与します。これらのプロセスは、組織の形成、創傷治癒、がん細胞の転移など、生体内での多くの重要な現象に不可欠です[3][4]。
● 疾患との関連
DblファミリーGEFの異常な活性化または発現は、がんを含む多くの疾患の発生に関連しています。特定のDblファミリーGEFが過剰に活性化されると、RhoファミリーGTPaseの制御が失われ、細胞増殖、運動性、および生存の異常な促進が引き起こされる可能性があります。このように、DblファミリーGEFは、がんの進行や転移の潜在的な治療標的として注目されています[3][4]。
● まとめ
Dblファミリーは、細胞内シグナル伝達において中心的な役割を果たすGEFであり、RhoファミリーGTPaseの活性化を通じて細胞の多様な機能を調節します。これらのタンパク質の正確な制御は、正常な細胞機能の維持に不可欠であり、その異常は疾患の発生に直接関連しています。Dblファミリーとその標的GTPaseの詳細な理解は、新たな治療戦略の開発に貢献する可能性があります。
- 参考文献・出典
-
[1] leading.lifesciencedb.jp/3-e001
[2] www.jstage.jst.go.jp/article/dermatol/108/12/108_1518/_pdf
[3] www.cosmobio.co.jp/product/detail/cytoskeleton_news_201605.asp?entry_id=16904
[4] www.cosmobio.co.jp/product/detail/product_g_p_1_20040625.asp?entry_id=5741
[5] www.jstage.jst.go.jp/article/kagakutoseibutsu1962/35/3/35_3_162/_pdf/-char/ja
[6] www.med.kobe-u.ac.jp/tougou/signal/Research.html
[7] nagoya.repo.nii.ac.jp/record/13916/files/k4913.pdf
[8] www1.gifu-u.ac.jp/~mb3_ulab/Research2.html
[9] lifesciencedb.jp/pne/Literature/get_pne_cgpdf.php?file=neRBr7DNNPLUS12Pk8hjmIs7A%3D%3D&number=4104&year=1996
[10] www.jsps.go.jp/file/storage/general/j-rftf/saishu_hyouka_08/02_life/p_pdf/l_04-06.pdf
Dblファミリーによる細胞の形態と運動の制御
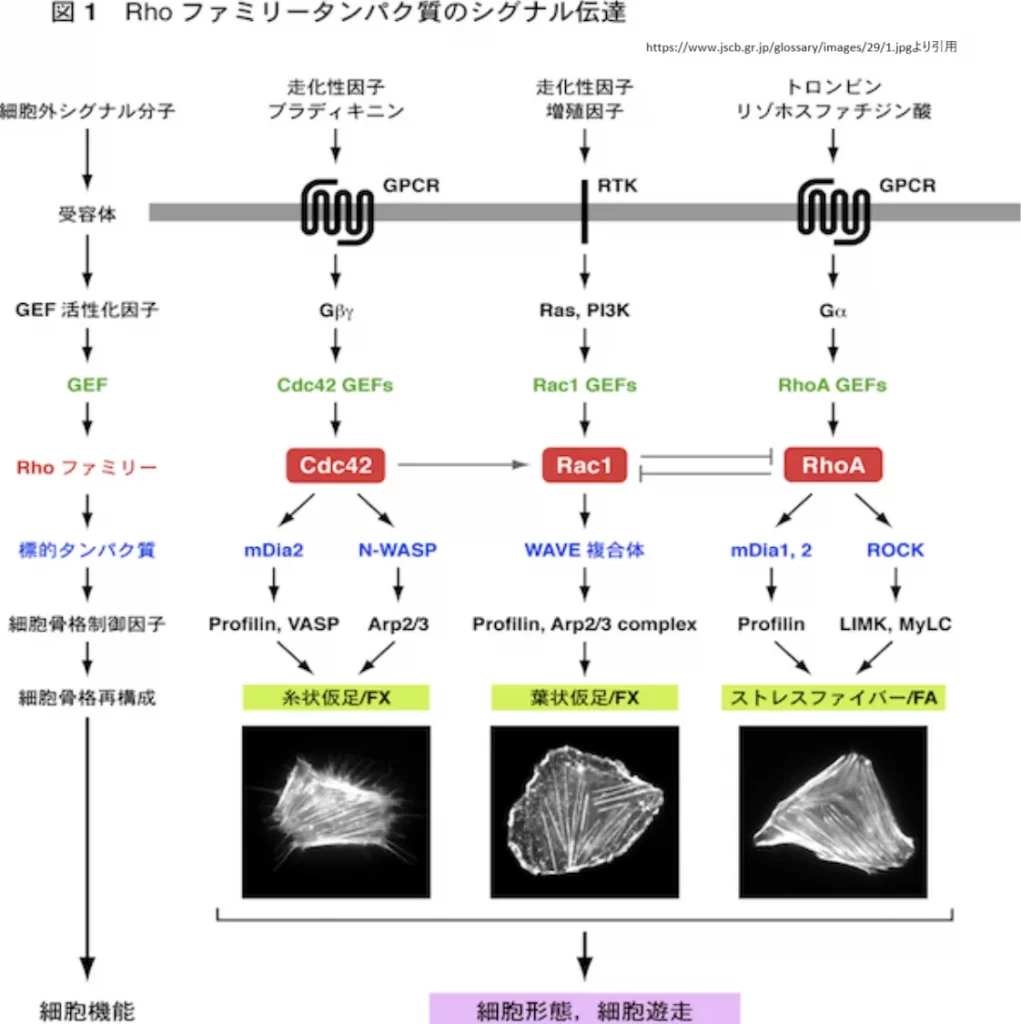
Dblファミリーは、細胞の形態と運動を制御する上で重要な役割を果たしています。このファミリーは、RhoファミリーGTP結合蛋白質(Rho、Rac、Cdc42)の活性を調節するグアニンヌクレオチド交換因子(GEF)で構成されています。RhoファミリーGTP結合蛋白質は、細胞形態、接着、運動などの制御を介して細胞の高次機能を調節し、その異常は細胞の癌化に深く関与していると考えられています[5]。
DblファミリーGEFは、細胞のアクチン細胞骨格系を制御することにより、細胞の形態変化や運動を促進します。例えば、Dblに関しては、Cdc42の標的分子であるACK-1チロシンキナーゼによりリン酸化され、その結果、Rho、Rac、Cdc42に対するGEF活性が上昇し、アクチン細胞骨格系が制御されることが見いだされました[5]。また、Ras-GRF-1に関しては、G蛋白質共役型レセプター刺激に応答し、G蛋白質βγサブユニットを介するシグナルによって、Ras-GRF-1がチロシンリン酸化され、Racに対するGEF活性が誘導されることが示されています[5]。
さらに、DblファミリーRhoGEFは、アミノ酸の一次構造からみる推定分子量が60kDa以上ある高分子蛋白質がほとんどであり、多くの場合、DH domainとPH domainを持つ以外にも、細胞の形態と運動の制御に関わる様々な機能を持っています[7]。
これらの研究結果から、Dblファミリーによる細胞の形態と運動の制御は、細胞内シグナル伝達の重要な部分を占めており、細胞の正常な機能維持だけでなく、病態の発生にも深く関わっていることが示されています。
- 参考文献・出典
-
[1] bsd.neuroinf.jp/w/index.php?mobileaction=toggle_view_desktop&title=Rho%E3%83%95%E3%82%A1%E3%83%9F%E3%83%AA%E3%83%BC%E4%BD%8E%E5%88%86%E5%AD%90%E9%87%8FG%E3%82%BF%E3%83%B3%E3%83%91%E3%82%AF%E8%B3%AA
[2] www.jstage.jst.go.jp/article/dermatol/108/12/108_1518/_pdf
[3] www.cosmobio.co.jp/product/detail/cytoskeleton_news_201605.asp?entry_id=16904
[4] www.cosmobio.co.jp/product/detail/product_g_p_1_20040625.asp?entry_id=5741
[5] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-12215044/
[6] webview.isho.jp/journal/detail/abs/10.11477/mf.2425200279
[7] www1.gifu-u.ac.jp/~mb3_ulab/Research2.html
[8] bibgraph.hpcr.jp/abst/pubmed/11738596?click_by=p_ref
[9] www.jsps.go.jp/file/storage/general/j-rftf/saishu_hyouka_08/02_life/p_pdf/l_04-06.pdf
[10] kaken.nii.ac.jp/en/report/KAKENHI-PROJECT-18J13111/18J131112018jisseki/
[11] seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2016.880416/data/index.html
第4章: 疾患とDblファミリー
Dblファミリーの異常が引き起こす疾患
Dblファミリーは、細胞内のシグナル伝達に関与する重要な分子群であり、特に細胞の形態変化、運動、増殖に関わっています。Dblファミリーの異常は、これらの細胞機能の調節失敗を引き起こし、様々な疾患の原因となり得ます。
1. 癌の形成: Dblファミリーの一員であるLBC腫瘍性タンパク質は、哺乳類の線維芽細胞において、小さなGTP結合タンパク質Rhoを特異的に活性化し、形質転換とアクチンストレス繊維形成を誘導します。この過程は、細胞のがん化に関連しています[1]。
2. 好中球機能異常: Dbl変異は、好中球の機能異常を引き起こし、リウマチ性疾患(RA)などの炎症性疾患の発症に関与している可能性があります。好中球は炎症反応において重要な役割を果たすため、その機能異常は炎症が終息しない原因となり得ます[5]。
3. 発達遅延: トリオのフレームシフト削除に関連する継承された世界的な発達遅延表現型についての研究があります。この研究では、知的障害、微小頭症、および顕著な発達遅延といった一貫した表現型が報告されています。トリオはDblファミリーに属する分子であり、このような異常が発達遅延の原因となることが示唆されています[6]。
4. ヌーナン症候群: PTPN11病的バリアントに関連する白血病のスペクトルは、ヌーナン症候群においても観察されます。ヌーナン症候群は、言語の遅れ、知覚運動障害、構音障害などの特徴を持つ遺伝性疾患です。PTPN11はDblファミリーに関連する分子であり、この症候群におけるその異常は、特定の白血病のリスクを高めることが示されています[4]。
これらの例から、Dblファミリーの異常が引き起こす疾患は多岐にわたり、細胞のシグナル伝達の調節失敗が様々な形で疾患に影響を及ぼすことがわかります。
- 参考文献・出典
-
[1] bibgraph.hpcr.jp/abst/pubmed/9030518?click_by=p_ref
[2] mhlw-grants.niph.go.jp/system/files/2017/172051/201711119A_upload/201711119A0007.pdf
[3] www.jpma.or.jp/information/evaluation/results/allotment/gbkspa0000000mw7-att/DS_202207_RWD.pdf
[4] grj.umin.jp/grj/noonan.htm
[5] www.jstage.jst.go.jp/article/naika1913/89/10/89_10_2054/_pdf
[6] bibgraph.hpcr.jp/abst/pubmed/27418539?click_by=p_ref
[7] www.jstage.jst.go.jp/article/jjrm1964/40/10/40_10_669/_pdf
[8] www.kanazawa-med.ac.jp/kenkyu/assets/H29houkokusyo.pdf
がんの発生におけるDblファミリーの関与
がんの発生においてDblファミリーは重要な役割を果たしています。Dblファミリーは、細胞内のシグナル伝達に関与する低分子量G蛋白質の活性を調節する因子であり、これらのG蛋白質は細胞の増殖、分化、運動などの多様な生物学的プロセスを制御しています。特に、RhoファミリーGTPaseの活性化に関与するDblファミリーのGEF(guanine nucleotide exchange factors)は、細胞の形態形成や運動性、細胞骨格の再編成など、がん細胞の浸潤や転移に必要な機能を調節することが知られています[1][2][4][7][8]。
DblファミリーのGEFは、RhoファミリーGTPaseを活性型のGTP結合型から不活性型のGDP結合型へと変換することで、これらのGTPaseの活性を制御します。この活性化は、細胞の成長因子に応答して行われることが多く、細胞の増殖や運動に直接影響を与えます。例えば、RhoGEFの一つであるDblは、細胞膜への局在化やエフェクタータンパク質との結合を通じてRhoファミリーGTPaseの活性化を促進し、細胞運動性や細胞骨格の調節に関与しています[2][3]。
また、Dblファミリーの異常は、がんの発生においても重要な役割を果たしています。例えば、Dblファミリーの一員であるPLEKHG2は、がんの浸潤や転移に関与する細胞運動の調節に関わっており、この遺伝子の異常はがんの進行に寄与する可能性があります[4]。さらに、Dblファミリー蛋白質は、EphA2受容体のシグナル伝達経路にも関与しており、上皮細胞の形態形成やがん化における役割が研究されています[3]。
総じて、Dblファミリーはがんの発生において、細胞の増殖や運動性を制御することで、がん細胞の浸潤や転移に重要な役割を果たしていると考えられます。これらの知見は、がん治療の新たな標的としてDblファミリーを考慮することの重要性を示唆しています。
- 参考文献・出典
-
[1] www.jstage.jst.go.jp/article/dermatol/108/12/108_1518/_pdf
[2] www.cosmobio.co.jp/product/detail/cytoskeleton_news_201605.asp?entry_id=16904
[3] repository.kulib.kyoto-u.ac.jp/dspace/bitstream/2433/199472/2/gseik00331.pdf
[4] www1.gifu-u.ac.jp/~mb3_ulab/Research2.html
[5] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-13214084/
[6] gakui.dl.itc.u-tokyo.ac.jp/cgi-bin/gazo.cgi?no=116250
[7] kaken.nii.ac.jp/ja/file/KAKENHI-PROJECT-23300349/23300349seika.pdf
[8] www.jstage.jst.go.jp/article/jsir2001/21/2/21_2_129/_pdf
第5章: 研究の最前線
最新の研究成果と進展
Dblファミリーのグアニンヌクレオチド交換因子(GEF)に関する最新の研究成果と進展は、細胞の高次機能の調節、特に細胞形態、接着、運動などにおけるRhoファミリーGTP結合蛋白質の役割の解明に焦点を当てています。これらの研究は、細胞の癌化に深く関与するRhoファミリー蛋白質の活性調節機構の理解を深めることを目的としています。
● Rhoファミリー蛋白質の活性調節機構
科学研究費助成事業(KAKEN)のプロジェクトによる研究では、DblファミリーGEFの中でも特にDblとRas-GRF-1に焦点を当て、リン酸化などのポストトランスレーショナル修飾がこれらのGEF活性に与える影響を検討しています。Dblに関しては、Cdc42の標的分子であるACK-1チロシンキナーゼによるリン酸化が、Rho、Rac、Cdc42に対するGEF活性を上昇させることが見いだされました。また、EGFレセプターがCdc42とアダプター分子Grb-2を介してACK-1を活性化し、Dblのリン酸化を通じてアクチン細胞骨格系が制御されることが明らかにされています[1]。
● 神経機能との関連
Rhoファミリーは神経突起形成・伸長の調節にも重要な役割を果たしており、精神遅滞などの遺伝性神経疾患の原因遺伝子としてRhoファミリーの情報伝達系に関わる分子が同定されています。これにより、神経回路形成におけるRhoファミリーの生理的役割の意義がクローズアップされています[2]。
● 神経発達障害発症機構の解明
日本神経化学会の研究では、Racシグナルの破綻による神経発達障害の発症機構が解明されています。PLEKHG2というDblファミリーのGEFが、マウスの大腳皮質神経細胞で発現しており、Rac3が中枢神経系に限局して発現していることが観察されました。これらの知見は、Rac機能の神経発生/発達における役割の理解を深めるものです[3]。
● 癌細胞の浸潤促進因子の構造生物学
上原記念生命科学財団の研究では、癌細胞の浸潤促進因子であるTiam1/2の構造生物学に関する研究が行われています。これらのGEFは、Rhoと直接結合してGDPの解離反応を促進するDbl Homology (DH) ドメインを持っており、癌細胞の浸潤や転移に関与していることが示唆されています[5]。
これらの研究成果は、細胞のシグナル伝達機構の理解を深めるだけでなく、癌や神経疾患などの病態の解明にも寄与しており、将来的な治療法の開発に向けた基盤を築いています。
- 参考文献・出典
-
[1] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-12215044/
[2] www.md.tsukuba.ac.jp/basic-med/molneurobiol/brain/kenkyu15n/kenkyu03.html
[3] neurochem-j.jp/10.11481/topics197/data/index.html
[4] seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2016.880416/data/index.html
[5] www.ueharazaidan.or.jp/houkokushu/Vol.29/pdf/144_report.pdf
[6] repository.dl.itc.u-tokyo.ac.jp/record/6491/files/A29992.pdf
[7] www.md.tsukuba.ac.jp/basic-med/molneurobiol/brain/kenkyu14n/kenkyu02.html
[8] pfwww.kek.jp/publications/pfnews/36_2/PFNews_36_2.pdf
[9] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-13216072/
[10] ja.wikipedia.org/wiki/%E3%82%B0%E3%82%A2%E3%83%8B%E3%83%B3%E3%83%8C%E3%82%AF%E3%83%AC%E3%82%AA%E3%83%81%E3%83%89%E4%BA%A4%E6%8F%9B%E5%9B%A0%E5%AD%90
[11] www.jstage.jst.go.jp/article/joma/130/3/130_109/_pdf/-char/ja
[12] repository.dl.itc.u-tokyo.ac.jp/record/2002529/files/A36178_review.pdf
[13] researchmap.jp/read0069583/presentations/13773240
日本におけるDblファミリー研究の現状
日本におけるDblファミリー研究の現状については、複数の研究機関や研究者が関わっていることが示されています。Dblファミリーは、細胞内のシグナル伝達において重要な役割を果たすRhoGEFタンパク質の一群であり、その構造や機能に関する研究が行われています。
まず、岐阜大学工学部化学・生命工学科では、三量体Gタンパク質Gβγサブユニットにより制御されるRho特異的グアニンヌクレオチド交換因子に関する研究が行われています。この研究では、RhoGEFタンパク質の構造から二つのファミリーの存在が知られており、その一つがDblファミリーであることが示されています[1]。
また、科学研究費補助金(KAKEN)のプロジェクトによる研究成果報告書では、RhoGEFタンパク質の全長またはマルチドメイン状態での立体構造解析と、GEFドメインと他の機能ドメインによる活性化制御機構の解明について報告されています[2]。この研究は、Dblファミリーを含むRhoGEFタンパク質の機能解明に貢献しています。
関節リウマチ疾患遺伝子としてのDblプロトオンコジーンに関する研究もあり、関節リウマチ(RA)患者検体におけるRhoファミリー活性化状況や、Dbl制御下での細胞内アクチン重合に起因する現象変化が検討されています[3]。この研究は、Dblファミリーが関節リウマチの発症機序に関与している可能性を示唆しています。
これらの研究成果は、日本におけるDblファミリー研究の現状を反映しており、細胞シグナル伝達の理解を深めるための基礎研究として、また疾患治療の新たなターゲットとしての可能性を探るための応用研究として、重要な役割を果たしています。
- 参考文献・出典
-
[1] seikagaku.jbsoc.or.jp/10.14952/SEIKAGAKU.2016.880416/data/index.html
[2] kaken.nii.ac.jp/ja/file/KAKENHI-PROJECT-15K06987/15K06987seika.pdf
[3] kaken.nii.ac.jp/ja/grant/KAKENHI-PROJECT-16790553/
[4] www.jstage.jst.go.jp/article/jkg/74/1/74_2/_pdf/-char/ja
[5] webview.isho.jp/journal/detail/abs/10.11477/mf.2425200279
[6] www.jstage.jst.go.jp/article/jsir2001/21/2/21_2_129/_pdf
[7] www1.gifu-u.ac.jp/~mb3_ulab/Research2.html
[8] www.hakatara.net/images/no3/3-7.pdf
Dbl family Rho GEFsに属する遺伝子
ABR
ALS2
ALS2CL
ARHGEF1
ARHGEF2
ARHGEF3
ARHGEF4
ARHGEF5
ARHGEF6
ARHGEF7
NET1
ARHGEF9
ARHGEF10
ARHGEF10L
ARHGEF11
ARHGEF12
AKAP13
MCF2L
ARHGEF15
ARHGEF16
ARHGEF17
ARHGEF18
ARHGEF19
MCF2
MCF2L2
TRIO
KALRN
ARHGEF25
ARHGEF26
NGEF
ARHGEF28
SPATA13
OBSCN
ECT2
ECT2L
ARHGEF33
ARHGEF34P
ARHGEF35
DNMBP
ARHGEF37
ARHGEF38
ARHGEF39
ARHGEF40
PLEKHG1
PLEKHG2
PLEKHG3
PLEKHG4
PLEKHG5
PLEKHG6
PLEKHG4B
BCR
FARP1
FARP2
FGD1
FGD2
FGD3
FGD4
FGD5
FGD6
ITSN1
ITSN2
PLEKHG7
PREX1
PREX2
RASGRF1
RASGRF2
SOS1
SOS2
TIAM1
TIAM2
VAV1
VAV2
VAV3

この記事の筆者:仲田洋美(医師)






